

Аденилоянтарная кислота
Ксантиловая кислота


 АТФ
АТФ
Фумаровая к-та
АМФ
ГМФ
Синтезу пиримидиновых нуклеотидов предшествует образование необычного азотистого основания - оротовой кислоты (ее нет в составе нуклеиновых кислот), содержащей пиримидиновое кольцо:
Источниками
атомов пиримидинового ядра являются:
глутамин, СО2
и аспарагиновая кислота
 СООН
СООН
Оротовая кислота
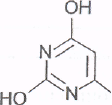
Синтезируется оротовая кислота из аммиака (NH2- глутамина), углекислого газа и аспарагиновой кислоты. Образовавшаяся оротовая кислота присоединяется к рибозо-5-фосфату и возникает пиримидиновый нуклеотид - ороидинмонофосфат. Далее оротовая кислота в составе этого нуклеотида преобразуется в обычные азотистые основания - тимин, урацил и цитозин, в результате чего появляются пиримидиновые нуклеотиды (тимидинмонофосфат - ТМФ, уридинмонофосфат - УМФ и цитидинмонофосфат - ЦМФ), входящие в состав нуклеиновых кислот.
В связи с такой важной ролью оротовой кислоты в синтезе нуклеотидов в спортивной практике в качестве пищевой добавки нередко применяется оротат калия.
Дезоксирибонуклеотиды (содержат дезоксирибозу) образуются из соответствующих рибонуклеотидов путем восстановления входящей в них рибозы в дезоксирибозу.
Синтез нуклеиновых кислот
Для синтеза нуклеиновых кислот используются мононуклеотиды обязательно в трифосфатной форме. Такие нуклеотиды содержат в своей молекуле три остатка фосфорной кислоты и обладают повышенным запасом энергии. Переход нуклеотидов в трифосфатную форму осуществляется путем взаимодействия с АТФ.
Для синтеза РНК необходимо четыре вида рибонуклеотидов (р-АТФ, р-ГТФ, р-УТФ и р-ЦТФ). Для синтеза ДНК используются дезоксирибонуклеотиды тоже четырех видов (д-АТФ, д-ГТФ, д-ТТФ и д-ЦТФ).
Синтез ДНК (репликация) интенсивно протекает во время клеточного деления. В процессе репликации в молекуле ДНК (родительская молекула) разрываются водородные связи между азотистыми основаниями обеих ее нитей, что приводит к раскручиванию двойной спирали
ДНК и образованию двух свободных нитей. К образовавшимся свободным нитям, как к матрицам, подходят нуклеотиды в трифосфатной форме и своими азотистыми основаниями с соблюдением принципа комплементарности (аденин - тимин и гуанин - цитозин) присоединяются к ним. Благодаря этому принципу создается нужная последовательность расположения нуклеотидов. По мере присоединения к матрице нуклеотиды связываются в полинуклеотидные нити, которые сразу же закручиваются с матрицей в двойную спираль. При этом от каждого нуклеотида отщепляется по два остатка фосфорной кислоты в форме дифосфата. В конечном счете, на каждой матрице возникает новая нить, которая по строению точно соответствует второй нити ДНК. В результате репликации синтезируются две новые молекулы ДНК (дочерние), которые являются точной копией родительской молекулы ДНК. В дочерних молекулах одна полинуклеотидная нить происходит из родительской молекулы, а другая синтезирована из нуклеотидов в процессе репликации.
Синтез РНК (транскрипция) также протекает с участием ДНК. В процессе транскрипции раскручивается только ограниченный участок ДНК и матрицей служит лишь одна освободившаяся нить ДНК. К этой нити, как к матрице, подходят нуклеотиды в трифосфатной форме, содержащие рибозу, и по принципу комплементарности располагаются в строго определенном порядке. Затем нуклеотиды соединяются в полинуклеотид, и от каждого из них отщепляется дифосфат. Образовавшаяся полинуклеотидная цепь с матрицей двойной спирали не образует и легко отходит от молекулы ДНК, после чего происходит восстановление ее двойной спирали. Таким образом происходит синтез информационных (иРНК), транспортных (тРНК) и рибосомных (рРНК) РНК.
Диффузия - самопроизвольное движение молекул из области высокой концентрации в направлении более низкой. Обусловлена диффузия тем, что все молекулы постоянно находятся в состоянии беспорядочного, хаотичного движения, Следствием диффузии является выравнивание концентрации.
Образовавшиеся α-кетокислоты далее подвергаются глубокому распаду и превращаются в конечные продукты СО2 и Н2О. Для каждой из 20 кетокислот (их образуется столько же, сколько имеется видов аминокислот) имеются свои специфические пути распада. Однако при распаде некоторых аминокислот в качестве промежуточного продукта образуется пировиноградная кислота, из которой возможен синтез глюкозы. Поэтому аминокислоты, из которых возникают такие кетокислоты, получили название глюкогенные. Другие же кетокислоты при своем распаде не образуют пирувата. Промежуточным продуктом у них является ацетилкоэнзим А, из которого невозможно получить глюкозу, но зато могут синтезироваться кетоновые тела. Аминокислоты, сооветствующие таким кетокислотам, называются кетогенные.
Второй продукт косвенного дезаминирования аминокислот - аммиак. Для организма аммиак является высоко токсичным. Поэтому в организме имеются молекулярные механизмы его обезвреживания.
По мере образования NН3 связывается во всех тканях с глутаминовой кислотой с образованием глутамина:
СООН О=С - NH2
I I
(СН2 )2 (СН2 )2
I + NH3+ АТФ I
СН - NН2 СН - NН2
I I
СООН СООН
Глутаминовая кислота Глутамин
или с аспарагиновой кислотой с образованием аспарагина.
Это временное обезвреживание аммиака. С током крови глутамин поступает в печень, где распадается опять на глутаминовую кислоту и NНз- Образовавшаяся глутаминовая кислота с кровью снова поступает в органы для обезвреживания новых порций аммиака. Освободившийся аммиак, а также углекислый газ в печени используются для синтеза мочевины.
Синтез мочевины - циклический, многостадийный процесс, потребляющий большое количество энергии. В синтезе мочевины очень важное участие принимает аминокислота орнитин. Эта аминокислота не входит в состав белков. Образуется орнитин из другой аминокислоты - аргинина, который присутствует в белках. В связи с важной ролью орнитина синтез мочевины получил название орнитиновый цикл.
В процессе синтеза к орнитину присоединяются две молекулы аммиака и молекула углекислого газа, и орнитин превращается в аргинин от которого сразу же отщепляется мочевина, и вновь образуется орнитин:
Н
карбамоилфосфат
орнитин
Н2N I О ~Р
цитруллин


Карбамоил-фосфат
 Фн
Фн