

Цис-АКОНИТАТ
ЦИТРАТ
ИЗОЦИТРАТ
α -КЕТОГЛУТАРАТ
АЦЕТИЛ Ко А
НАД+
НАДН2
ОКСАЛО-АЦЕТАТ
СУКЦИНИЛ-Ко А
МАЛАТ
Н3РО4
+ ГДФ
ГТФ
HS-Ko A
Е - ФАД
Е-ФАДН2
ФУМАРАТ
Н2О
Н2О
НАДН2
НАД+
Н2О
НАД+
НАДН2
СО2
HS-Ko A
CO2
HS-Ko A
Н2ОСукцинат


















ЦТК
Обмен липидов.
Поскольку в пище присутствуют разнообразные жиры, то в результате их переваривания образуется большое количество разновидностей жирных кислот. Продукты расщепления жира всасываются слизистой тонкой кишки. Глицерин растворим в воде, поэтому его всасывание происходит легко. Жирные кислоты, нерастворимые в воде, всасываются в виде комплексов с желчными кислотами. В клетках тонкой кишки глицерин вновь соединяется с жирными кислотами, но только с теми, которые входят в состав жиров организма человека. В результате: синтезируется человеческий жир.
Вновь образовавшийся (ресинтезированный) жир по лимфатическим сосудам, минуя печень, поступает в большой круг кровообращения и откладывается в запас в жировых депо. Главные жировые депо организма - подкожная жировая клетчатка, большой и малый сальники, околопочечная капсула.
Катаболизм жиров
Использование жира в качестве источника энергии начинается с его выхода из жировых депо в кровяное русло. Этот процесс называется мобилизацией жира. Мобилизация жира ускоряется под влиянием симпатической нервной системы и гормона адреналина.
Основные превращения жира происходят в печени, где имеются активные ферменты жирового обмена.
В печени жир прежде всего подвергается гидролизу и превращается, так же как и в кишечнике, в глицерин и жирные кислоты.
Образовавшийся глицерин легко переходит в фосфоглицериновый альдегид, который является также промежуточным продуктол распада углеводов и поэтому вовлекается в углеводный обмен.
Жирные кислоты, являясь веществами химически неактивными, вначале активируются с использованием энергии АТФ и связываются своим переносчиком - коферментом А (комплекс жирная кислота кофермент А называется ацилкофермент А):
АТФ + R— СООН + НS-КоА —> R — СО ~ SКоА
Жирная -АМФ
кислота — ФФ Ацил-коэнзим А
ацил-ko А-синтетаза
Образовавшийся ацилкофермент А далее с помощью еще одного переносчика – l- карнитина поступает в митохондрии, где происходит окисление жирной кислоты.
Окисление жирных кислот протекает в два этапа. На первом этапе, называемом β-окислением (данное название обусловлено тем, что окислению подвергается углеродный атом жирной кислоты, находящийся в (β-положении), от жирной кислоты, связанной с коферментом А, дважды отщепляется по два атома водорода, которые затем по дыхательной цепи передаются на молекулярный кислород. В итоге образуется вода, и за счет выделяющейся при этом энергии осуществляется синтез пяти молекул АТФ. Завершается β-окисление отщеплением от жирной кислоты двууглеродного фрагмента в виде ацетилкофермента А:
Получается Укороченная жирная кислота
β-окисление многократно повторяется до тех пор, пока жирная кислота полностью не превратится в ацетил-КоА, количество молекул которого равно половине числа атомов углерода в исходной жирной кислоте. Как уже отмечалось, каждое отщепление ацетилкофермента А сопровождается синтезом пяти молекул АТФ, осуществляемым тканевым дыханием.
Вторым этапом окисления жирных кислот является цикл трикарбоновых кислот (ЦТК) или цикл Кребса, в котором происходит дальнейшее окисление остатка уксусной кислоты, входящей в ацетилкофермент А, до углекислого газа и воды. При окислении одной молекулы ацетилкофермента А выделяется 12 молекул АТФ. В целом окисление жирных кислот до СО2 и Н2О дает большое количество энергии. Например, в случае окисления пальмитиновой кислоты (С15Н31СООН) семь раз протекает β-окисление, что приводит к образованию 35 молекул АТФ и 8 молекул ацетилкофермента А. При дальнейшем окислении 8 молекул ацетилкофермента А в цикле Кребса еще синтезируется 96 молекул АТФ. Вычтя из полученной суммы молекул АТФ одну молекулу, энергия которой была затрачена на активацию жирной кислоты, получаем окончательный результат; при окислении молекулы пальмитиновой кислоты образуется 130 молекул АТФ.
Окисление жира протекает в митохондриях при обязательном использовании молекулярного кислорода, что существенно ограничивает скорость этого процесса. Поэтому за счет окисления жиров можно обеспечить энергией только работу средней мощности, но зато очень продолжительную, так как запасы жира в организме весьма значительны. При избыточном образовании ацетил-кофермента А, что обычно бывает в печени, вместо цикла Кребса происходит реакция конденсации. В результате конденсации остатки уксусной кислоты, соединяясь попарно, превращаются в кетоновые тела, а кофермент А выделяется в свободном виде:
СН3СО~S-ko A + CH3СO~S-ko A CH3-CO-CH2CO~S-koА+ HS-koA
Ацетоацетил-Ко А
Ацетоацетил-КоА-тиолаза
1) CH3-CO-CH2CO~S-koА +НАД-Н2 СН3-СНОН-СН2-СООН +НАД
Ацетоацетил-Ко А β-оксимасляная к-та
D-β-гидроксибутиратдегидрогеназа
2) CH3-CO-CH2CO~S-koА + Н2О CH3-CO-CH2-COОН + HS-koA
Ацетоацетил-Ко А Ацетоуксусная к-та
Ацето-ацетил-КоА-гидролаза (деацилаза)
1) и 2) – образование Кетоновых тел (ацетоуксусной к-ты СН3СОСН2СООН, β-гидроксимасляной кислоты СН3СНОНСН2СООН и ацетона СН3СОСН3).
С током крови кетоновые тела поступают во все ткани. Однако большая часть кетоновых тел извлекается из крови органами, имеющими высокие энергозатраты; миокардом, скелетными мышцами, почками. В этих органах с участием их собственного кофермента А кетоновые тела вновь переходят в ацетилкофермент А:
D-β-гидроксибутиратдегидрогеназа
СН3-СНОН-СН2-СООН +НАД CH3-CO-CH2-COОН +НАДН2
β- гидроксимасляная к-та Ацетоуксусная кислота
Ацил-КоА-синтетаза
СН3-СНОН-СН2-СООН + АТФ+ HS-koA CH3-CO-CH2CO~S-koА + АМФ
Ацетоуксусная к-та +Пирофосфат
Ацетоацетил-Ко А
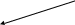
+ HS-koA
2СН3-СО~ S-koA
Ацетил-КоА Ацето-ацетил-КоА-гидролаза
Далее ацетилкофермент А, окисляясь в цикле Кребса, дает этим органам необходимую энергию для их функционирования. Особенно велика роль кетоновых тел при обеспечении энергией продолжительных физических нагрузок. При накоплении кетоновых тел в крови возможно образование ацетона.
СН3-СНОН-СН2-СООН -------- СН3-СО- СН3 +CO2
Ацетоуксусная кислота АЦЕТОН
Это вещество в организме не используется и выделяется легкими и почками.