

Биологическое окисление
Как уже отмечалось, в процессе обмена веществ в организме происходят различные реакции окисления, которые объединяют термином биологическое окисление.
Основным типом биологического окисления является тканевое дыхание, протекающее в клеточных митохондриях (в связи с этим тканевое дыхание еще называется митохондриальным окислением).
Тканевое дыхание
Тканевое дыхание - это основной способ получения АТФ, используемый всеми клетками организма (кроме красных клеток крови).
В процессе тканевого дыхания от окисляемого вещества отнимаются два атома водорода (два протона и два электрона) и по дыхательной цепи, состоящей из ферментов и коферментов, передаются на молекулярный кислород - О2) доставляемый кровью из воздуха во все ткани организма. В результате присоединения атомов водорода к кислороду образуется вода. За счет энергии, выделяющейся при движении электронов по дыхательной цепи, в митохондриях осуществляется синтез АТФ из АДФ и фосфорной кислоты. Обычно образование одной молекулы воды сопровождается синтезом трех молекул АТФ.
В упрощенном виде тканевое дыхание может быть представлено следующей схемой:
АН2 + 1/2 О2 А + Н2О
Окисленное
вещество

3 АДФ + 3 Н3РО4 ЗАТФ
В качестве субстратов окисления (т. е. веществ, от которых отнимается водород) в тканевом дыхании используются разнообразные промежуточные продукты распада белков, углеводов и жиров. Однако наиболее часто окислению подвергаются промежуточные продукты цикла трикарбоновых кислот (ЦТК) - цикла Кребса (изолимонная, а-кетоглутаровая, янтарная и яблочная кислоты). Цикл Кребса - это завершающий этап катаболизма, в ходе которого происходит окисление остатка уксусной кислоты, входящей в ацетилкофермент А, до СО2 и Н2О.
В свою очередь, ацетилкофермент А - это универсальный метаболит организма, в который при своем распаде превращаются главные органические вещества - белки, углеводы и жиры.
Тканевое дыхание представляет собой сложный ферментативный процесс.
Все ферменты тканевого дыхания делятся на три группы:
никотинамидные дегидрогеназы,
флавиновые дегидрогеназы и
цитохромы.
Никотинамидные дегидрогеназы отнимают два атома водорода от окисляемого субстрата и временно присоединяют их к своему коферменту НАД (никотинамидадениндинуклеотид). По строению НАД является динуклеотидом, в котором два нуклеотида соединяются между собой остатками фосфорной кислоты. В состав одного из нуклеотидов в качестве азотистого основания входит амид никотиновой кислоты (никотинамид, витамин РР), вторым нуклеотидом является аденозинмонофосфат (АМФ):

Отнимаемые от окисляемого вещества атомы водорода присоединяются непосредственно к никотинамиду (отсюда название данных ферментов - никотинамидные дегидрогеназы), при этом НАД переходит в свою восстановленную форму НАД Н2:
АН2 + НАД А + НАД-Н2
Окисляемое вещество
Флавиновые дегидрогеназы отщепляют два атома водорода от образовавшегося НАД-Н2 и временно присоединяют их к своему кофер-менту ФМН (флавинмононуклеотид). По строению этот кофермент является мононуклеотидом, содержащим витамин В2 (рибофлавин):
Флавин
(изоаллоксазин)
Рибитол
Фосфат


Рибофлавин (витамин В2)
Два атома водорода, отнимаемые флавиновыми дегидрогеназами от восстановленного НАД (НАД-Н2), присоединяются к флавину, выполняющему роль акцептора водорода. В результате этой стадии образуется восстановленная форма кофермента - ФМН-Н2:
НАД-Н2 + ФМН — —> НАД + ФМН-Н2
В некоторых случаях флавиновые дегидрогеназы, подобно никоти-намидным, отнимают два атома водорода непосредственно от окисляе-
мых веществ. Такие флавиновые дегидрогеназы используют кофермент
ФАД (флавинадениндинуклеотид), похожий по строению на ФМН и тоже содержащий витамин В2:
АН2 + ФАД А + ФАД- Н2
Окисляемое вещество Окисленное вещество
Следующая группа ферментов - цитохромы. Эти ферменты участвуют только в переносе электронов. По строению цитохромы похожи на одну из субъединиц гемоглобина. Молекула цитохрома состоит из полипептида и гема. Но в отличие от гемоглобина железо, входящее в гем цитохромов, имеет переменную валентность. Способность железа обратимо переходить из окисленной формы в восстановленную
(Fе3+
+ е
Fе2+)
обеспечивает возможность переноса
электронов данными
ферментами.
С помощью цитохромов (их имеется несколько: b, с, а, a3) электроны от восстановленных коферментов ФМН-Н2 и ФАД-Н2 передаются на молекулярный кислород (О2), который при этом переходит в активную, анионную, форму - О2~. Далее активный кислород (О2~) связывается с ионами водорода (протонами), которые тоже отщепляются от ФМН-Н2 или от ФАД-Н2. Присоединение ионов водорода к аниону кислорода приводят к образованию воды.
Таким образом, на всем протяжении дыхательной цепи наблюдается передвижение электронов. Движение электронов вызвано тем, что все участники дыхательной цепи располагаются по мере возрастания их окислительно-восстановительных потенциалов.
Окислительно-восстановительный потенциал, или редокс-потенциал, характеризует способность вещества принимать и удерживать электроны. Поэтому электроны переносятся от вещества с низким редокс-потенциалом к веществу с более высоким редокс-потенциалом.
Поскольку самое низкое значение редокс-потенциала имеет окисляемое вещество, а самое высокое - кислород, то в итоге электроны от окисляемого вещества поступают на молекулу кислорода.
Как уже отмечалось, движение электронов по дыхательной цепи со- провождается выделением энергии. Около половины энергии движения, электронов аккумулируется в макроэргических связях молекул АТФ. Другая часть энергии выделяется в виде тепла. Синтез АТФ происходит при переносе электронов с НАД-Н2 на ФМН, с цитохрома b на цитохром с и c цитохрома а на цитохром а3. Всего при переносе двух атомов водорода на кислород (г. е. в расчете на одну образовавшуюся молекулу воды) синтезируется три молекулы АТФ.
Некоторые субстраты (жирные кислоты, янтарная кислота и др.) имеют более высокий редокс-потенциал, чем НАД. Поэтому они не могут окисляться ншкотинамидными дегидрогеназами. В этом случае отнятие атомов водорода от таких субстратов осуществляется флавиновыми дегидрогеназами. Из-за отсутствия никотинамидных дегидрогеназ при окислении таких веществ образуется только две молекулы АТФ.
На рис. представлена схема тканевого дыхания, включающая все группы ферментов.
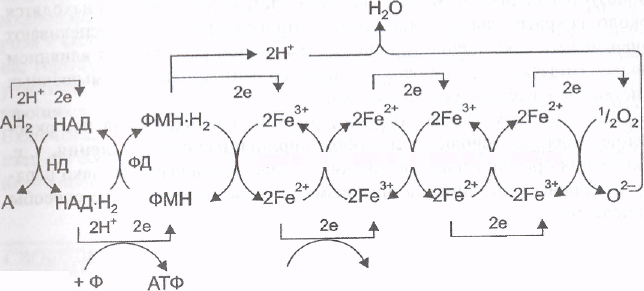

АДФ
Рис. Схема
тканевого дыхания
Образование АТФ в процессе тканевого дыхания часто обозначается терминами: окислительное фосфорилирование, дыхательное фосфорилирование, аэробное фосфорилирование, или аэробный синтез АТФ.
В сутки в организме за счет тканевого дыхания возникает не менее 40 кг АТФ, а у спортсменов еще больше. Поэтому этот процесс потребляет большое количество окисляемых веществ и кислорода.
При незначительной потребности клеток в АТФ тканевое дыхание протекает с низкой скоростью. Если клетка начинает использовать большое количество АТФ, то скорость тканевого дыхания возрастает и может достигнуть максимальных величин. Такой характер изменения скорости обусловлен тем, что активатором ферментов тканевого дыхания является избыток АДФ, который возникает в клетке только при интенсивном использовании АТФ.
Митохондрии, в которых протекает тканевое дыхание, имеются во всех клетках (кроме красных клеток крови) и представляют собою вытянутые палочковидные образования длиной 2-3 мкм* и толщиной около 1 мкм. Количество митохондрий в клетках может достигать тысячи и более. Митохондрии снаружи окружены двойной мембраной. Внешняя мембрана гладкая, а внутренняя складчатая, с большой поверхностью Ферменты тканевого дыхания встроены во внутреннюю мембрану и располагаются в ней в виде отдельных скоплений, называемых дыхательными ансамблями. Каждый дыхательный ансамбль содержит все необходимые ферменты для обеспечения переноса электронов в процессе тканевого дыхания. Благодаря строго упорядоченному расположению ферментов в дыхательных ансамблях передвижение электронов по дыхательной цепи осуществляется с большой скоростью.
В клетках митохондрии часто располагаются в том месте, где используется энергия АТФ. В мышечных клетках митохондрии находятся около сократительных элементов - миофибрилл - и обеспечивают энергией их сокращение в процессе мышечной работы. Под влиянием систематических тренировок количество митохондрий в мышечных клетках значительно увеличивается.
Как выше отмечалось, тканевое дыхание (митохондриальное окисление) является основным способом биологического окисления, т. е. окисления органических соединений в живом организме. Однако наряду с тканевым дыханием в организме еще имеются и другие способы окисления.