

Міністерство науки та освіти, молоді та спорту
Дніпропетровський національний університет імені Олеся Гончара
Центр заочної та дистанційної освіти
Кафедра мікробіології та вірусології
КОНТРОЛЬНА РОБОТА
з медичної мікробіології за питаннями:
1. Характеристика бактерій роду Salmonella
2. Загальна характеристика родини Enterobacteriaceae
Виконав:
студентка групи ББ-09-2з Донець Н.М.
Перевірив:
к.б.н., доцент Гаврилюк В.Г.
Дніпропетровськ
2012 р.
Содержание
Вступление
1. Характеристика бактерий рода Salmonella
1.1. Биологические свойства
1.2. Патогенез
1.3. Клинические проявления заболеваний
1.4. Иммунитет
1.5. Лабораторная диагностика
1.6. Лечение и профилактика сальмонеллёзов
2. Общая характеристика семейства Enterobakteriaceae
2.1.Биологические свойства энтеробактерий
2.2. Классификация энтеробактерий
2.2.1. Род Escherihia
2.2.2. Род Shigella
2.2.3. Род Salmonella
2.2.4. Род Yersinia
Список литературы
Вступление
Согласно второму изданию руководства "Bergey's Manual of Systematic Bacteriology (2005) бактерии семейства Enterobacteriaceae входят в домен Bacteria, тип Proteobacteria, класс Gammaproteobacteria, порядок Enterobacteriales. По состоянию на 01.06.2011 семейство Enterobacteriaceae включает 47 родов (табл. 1)
Таксономическое определение семейства Enterobacteriaceae, представленное в руководстве "Bergey's Manual of Systematic Bacteriology" (1984), также сохраняет значение. "Семейство Enterobacteriaceae - это грамотрицательные тонкие палочки 0,3-1,0 х 0,6-1,0 мкм, подвижные перитрихи (кроме Tatumella) или неподвижные. Они не образуют эндоспоры или микроцисты, не кислотоустойчивы. Растут в присутствии кислорода или без него. Хорошо развиваются в пептонных и мясных средах, обычно на среде Мак Конки. Некоторые используют D-глюкозу как единственный источник углерода, другие требуют в качестве добавок витамины и (или) аминокислоты. Питание осуществляют за счет органических соединений; механизм метаболизма дыхательный и ферментативный. Не галлофилы. При ферментации D-глюкозы, других углеводов и многоатомных спиртов продуцируют кислоты и видимые объемы газа. Каталазоположительны, за исключениемShigella dysenteriae серовара 1 и Xenorhabdus nematophilus, оксидазонегативны. Нитраты редуцируют в нитриты, за исключением некоторых штаммов Erwinia и Yersinia. Содержание Г+Ц в ДНК 38-60 моль %. ДНК видов, принадлежащих к большинству родов, родственны между собой не более чем на 20 %. Такая же степень родства и к Escherichia coli - типовому виду всего семейства. Исключение составляют некоторые виды Yersinia, Proteus, Providencia, Hafnia, Edwardsiella, ДНК которых имеют 10 - 20 % родства с ДНК видов, принадлежащих к другим родам. Все изученные виды содержат энтеробактериальный общий антиген (СА), кроме Erwinia chrysanthemi. Типовой род - Escherichia Castellani и Chalmere 1919 - определен Юридической комиссией Международного комитета систематической бактериологии в 1958 году". Следует отметить, что бактерии Plesiomonas shigelloides, включенные в семейство энтеробактерий, имеют оксидазу; бактерии Serratia marcescens subsp. sakuensis имеют эндоспоры.
Энтеробактерии — возбудители кишечных инфекций человека относятся исключительно к родам escherichia, shigella, salmonella и yersinia.
1. Характеристика бактерий рода Salmonella
Бактерии рода Salmonella представляют обширную группу микроорганизмов, включающую более 1600 серологических типов. Значение серологических типов сальмонелл в патологии человека различно. Сказанное относится к энтеропатогенным кишечным палочкам второй категории, обладающим поверхностным соматическим К-антигеном В или L, препятствующим О-агглютинации и инактивирующимся прогреванием в течение 1 ч. Энтеропатогенные кишечные палочки «Крым» не имеют К-антигена и поэтому агглютинируются в живом и убитом состоянии гомологичной сывороткой до ее предельного титра. Хозяевами подавляющего большинства других типов сальмонелл являются различные представители домашних и диких животных, птицы, рыбы, рептилии. Однако, будучи полипатогенными микроорганизмами, многие из этих бактерий опасны не только для животных, но и для людей.
1.1.Биологические свойства
Морфологически все бактерии, относящиеся к роду сальмонелл, представляют грамотрицательные палочки (длиной 2—4 мкм), не образующие спор. Подавляющее большинство из них подвижно благодаря наличию перитрихиально расположенных жгутиков (рис. 1).
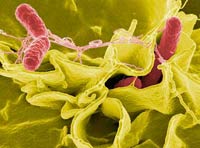
Рис. 1. Микрофотография бактерий рода Salmonella
Характеризуются следующими ферментативными свойствами: не разлагают лактозу, сахарозу, адонит, салицин, не разжижают желатин, не образуют индола, не расщепляют мочевину; разлагают глюкозу и маннит с образованиемгаза. Большинство сальмонелл продуцирует сероводород. Из числа аминокислот способны декарбоксилировать лизин, многие — аргинин и орнитин, но не способны декарбоксилировать глутаминовую кислоту. Хотя ферментативные свойства сальмонелл не всегда стабильны, культурально-биохимический метод типирования является необходимым в разграничении серологических типов, имеющих одинаковую антигенную структуру.
Антигены. Сальмонеллы имеют сложную антигенную структуру.Они содержат О-антигены (соматические) и Н-антигены (жгутиковые). Некоторые сальмонеллы имеют К-антиген.Сальмонеллы брюшного тифа наряду с О- и Н-антигенами могут содержать Vi-антиген. Он относится к К-антигенам и представляет собой полимер N-ацетилгалактозаминоуроновой кислоты. Наличие Vi-антигена препятствует агглютинации бактерий О-сыворотками. Видовая идентификация тифопаратифозной группы бактерий основана на изучении ферментативных свойств и обнаружении видо- и типоспецифических антигенов. Все представители рода сальмонелл различаются между собой двумя основными антигенами: термостабильным соматическим О-антигеном и термолабильным жгутиковым Н-антигеном. Жгутиковый Н-антиген у подавляющего большинства сальмонелл имеет две фазы: I — специфическую и II — неспецифическую. Структура антигенного аппарата, детально изученная Кауфманом и Уайтом, легла в основу современной серологической классификации сальмонелл.
Согласно классификации Кауфмана и Уайта, все многочисленные представители рода сальмонелл по О-антигену объединены в группы А, В, С, Д, Е и ряд редких типов (М); каждая из групп включает различные серологические типы,устанавливаемые на основании Н-антигенов.
Ежегодно схема Кауфмана — Уайта пополняется 40 — 80 новыми серотипами сальмонелл, описываемыми в разных странах и вводимыми в схему после углубленного изучения их в Международном центре по сальмонеллезам. Как уже указывалось выше, сейчас идентифицировано свыше 2 тыс. различных сальмонелл. Однако в патологии у людей имеют широкое распространение и играют эпидемиологическую роль 40 — 50 из них, причем чаще других S. typhimurium, S. Heidelberg, S. derby, S. anatum, S. panama. В последние годы, кроме биохимического и серологического методов типирования сальмонелл, используют также метод фаготипирования их.
Выявленный фаготип, особенно в пределах одного серотипа (чаще S. typhimurium), становится как бы «меткой», позволяющей более точно проводить эпидемиологический анализ, как отдельных заболеваний, так и эпидемических вспышек. Используется, как международный набор фагов [Felix и Callow), так и полученный в Тбилисском НИИВС (И. Г. Чиракадзе и Т. Т. Чанишвили). В настоящее время широкое распространение получил особый биологическй вариант S. typhimurium, в пределах которого установлено наличие трех основных биологических вариантов (биоваров), идентичных по антигенной структуре, но четко дифференцируемых по патогенности для белых мышей в опытах энтеральногозаражения. Биовар I — типичные культуры S. typhimurium, высокопатогенные для мышей. Биовар IIS менее патогенен для мышей, чувствителен к антибиотикам, описан в Ленинграде в 1950 г. как возбудитель внутрибольничной инфекции среди новорожденных и детей раннего возраста. Биовар IIR, так называемый «госпитальный штамм», также мало патогенен для мышей, но резистентен к антибиотикам (за исключением гентамицина и полимиксина М), не способен ферментировать α-тартрат. В последние годы биовар IIR получил широкое распространение как возбудитель внутрибольничных инфекций в детских стационарах, хотя может циркулировать и вне больничных очагов, преимущественно у детей 1-го года жизни.
Бактерии, входящие в состав каждой серологической группы, по совокупности I и II фаз Н-антигенов в свою очередь подразделяются на серологические типы. Установление серологической группы и серологического типа сальмонелл основано на определении формулы их антигенного строения по идентификации рецепторов соматических О-антигенов и жгутиковых Н-антигенов I и II фаз в реакции агглютинации с моноре-цепторными сыворотками. В настоящее время род сальмонелл по рекомендации Международного номенклатурного комитета подразделен на 4 вида (табл. 31). Вид I, обозначенный S. kauffmani, объединяет большую часть патогенных для человека сальмонелл серологических групп А, В, С, D, Е. Ко II виду, S. salamal, отнесены представители, отличающиеся от I вида некоторыми биохимическими признаками, в частности способностью разжижать желатин и расщеплять малонат натрия. Вид III, S. arisona, отличается от двух предыдущих видов способностью ферментировать (большей частью замедленно).
У человека S. arisona обнаруживаются при лихорадочных состояниях с явлениями диареи и гастроэнтерита. В последние годы описания заболеваний людей, причиной возникновения которых авторы считают бактерии Arisona, значительно участились. К IV виду, S. houtenae, отнесены изъятые из II вида «атипичные» в биохимическом отношении сальмонеллы, ферментирующие салицин и растущие в присутствии цианида калия. Чувствительность к цианиду калия отражает, по-видимому, определенную общебиологическую закономерность: все патогенные для человека кишечные бактерии чувствительны к цианиду калия и не растут в его присутствии (шигеллы, сальмонеллы); все непатогенные виды кишечных бактерий способны расти на средах с цианидом калия. Бактерии рода Escherichia и Arisona, включающие патогенных для человека представителей, подобно шигеллам и сальмонеллам чувствительны к этому яду.
Факторы патогенности.
1.Факторы адгезии и колонизации.
2. Способность к внутриклеточному паразитированию, препятствовать фагоцитозу, размножаться в клетках лимфоидной ткани выражены у возбудителей брюшного тифа, паратифов-А и -В, способствуя хроническому носительству.
3.Эндотоксин (ЛПС).
4. Термолабильные и термостабильные энтеротоксины.
5. Цитотоксины.
6. Существенное значение имеют плазмиды вирулентности и R- плазмиды.
7. Vi - антиген ингибирует действие сывороточных и фагоцитарных бактериоцидных факторов.
Основными факторами патогенности сальмонелл является их способность проникать в макрофаги и размножаться в лимфоидных образованиях собственно слизистого слоя тонкого кишечника (пейеровы бляшки, солитарные фолликулы), а также продукция эндотоксина.
Экология и эпидемиология. Распространены повсеместно. Подавляющее большинство сальмонелл, циркулирующих во всех странах, так же как и у нас, относится к четырем серологическим группам — В, С, D, Е (98%) и только 2% — к остальным серогруппам. Почти 2/3 заболеваний связано с сальмонеллами группы В (S. typhimurium, S. heidelberg, S. derby). В нашей стране обязательная повсеместная адресная регистрация всех сальмонеллезов (не только пищевых токсикоинфекций, но и отдельных разрозненных спорадических случаев) введена с 1959 г. В связи с этим суждение о распространенности сальмонеллезов в предшествующие годы крайне затруднено.
Имеются сведения о росте сальмонеллезных заболеваний в ряде стран Европы и США в период второй мировой войны и послевоенные годы, в основном за счет групповых вспышек. Аналогичная картина наблюдалась и у нас. В последнее 20-летие в нашей стране отмечается рост сальмонеллезных заболеваний, особенно у детей, в основном за счет спорадических случаев, которые составляют до 90% в сумме всех сальмонеллезов [Килессо В. А., 1980]. Однако и современный учет сальмонеллезных заболеваний, по-видимому, неполный. Это связано с трудностью клинической диагностики, наличием легких, стертых и бессимптомных форм болезни. Поэтому в действительности фактическая заболеваемость сальмонеллезом выше регистрируемой. Представляют интерес данные, полученные М. Г. Чахутинской (1968) при сравнительном анализе клинических проявлений болезни и лабораторных данных при разрозненных и групповых заболеваниях сальмонеллезом. Исследования показали, что высеваемость сальмонелл в обеих группах была идентична (по систематичности, количественной характеристике, длительности). Это позволило автору прийти к заключению о том, что между групповыми и спорадическими заболеваниями принципиальной разницы нет. Подтверждением данного положения является и то, что в 7 — 12% наблюдений спорадических сальмонеллезов удается выявить небольшие семейные вспышки. Наряду с общим подъемом уровня заболеваемости сальмонеллезом, в последнее время отмечается и неуклонное увеличение числа циркулирующих серотипов возбудителей этой инфекции. Если в начале настоящего столетия было выделено только 10 серологических типов сальмонелл, в 1934 — 1944 годах — 44 серотипа, то, по данным Всесоюзного центра по сальмонеллезам, за период с 1968 по 1972 г. зарегистрировано 334 серологических типа сальмонелл, и в настоящее время их выделено уже 500 [Килессо В. А., 1980]. Процесс возрастания числа обнаруживаемых серотипов сальмонелл является характерным для всех стран. В США в 1968 г. циркулировали 154 серотипа сальмонелл, а к 1970 г. число их возросло до 241 [Aserkoff et al., 1970]. В Париже с 1970 по 1972 г. Национальным центром изучения сальмонелл было идентифицировано 383 серотипа, из них 25, ранее не известных [Le Minor L., Le Minor S., 1973].