

2.11. Влияние внешних и внутренних факторов на интенсивность фотосинтеза
В физиологии растений пользуются двумя понятиями: истинный и наблюдаемый фотосинтез. Это обусловлено следующими соображениями. Скорость или интенсивность фотосинтеза характеризуется количеством СО2, поглощенного единицей поверхности листа в единицу времени. Определение интенсивности фотосинтеза проводят газометрическим методом по изменению (уменьшению) количества СО2 в замкнутой камере с листом. Однако, вместе с фотосинтезом идет процесс дыхания, во время которого выделяется СО2. Поэтому получаемые результаты дают представление об интенсивности наблюдаемого фотосинтеза. Для получения величины истинного фотосинтеза необходимо сделать поправку на дыхание. Поэтому перед опытом определяют в темноте интенсивность дыхания, а потом уже интенсивность наблюдаемого фотосинтеза. Затем количество СО2, выделенного при дыхании, прибавляют к количеству СО2, поглощенного на свету. Внося эту поправку, считают, что интенсивность дыхания на свету и в темноте одинакова. Но эти поправки не могут дать оценку истинного фотосинтеза потому, что, во-первых, при затемнении листа исключается не только истинный фотосинтез, но и фотодыхание; во-вторых, так называемое темновое дыхание в действительности зависит от света (см. дальше).
Поэтому во всех экспериментальных работах по фотосинтетическому газообмену листа отдают преимущество данным по наблюдаемому фотосинтезу. Более точный метод изучения интенсивности фотосинтеза – метод меченных атомов (измеряют количество поглощенного 14СО2).
В том случае, когда пересчет количества поглощенного СО2 на единицу поверхности трудно провести (хвойные, семена, плоды, стебель), полученные данные относят к единице массы. Учитывая, что фотосинтетический коэффициент (отношение объема выделенного кислорода к объему поглощенного СО2 равен единице, скорость наблюдаемого фотосинтеза можно оценивать по количеству миллилитров кислорода, выделенной единицей площади листа за 1 час.
Для характеристики фотосинтеза пользуются и другими показателями: квантовый расход, квантовый выход фотосинтеза, ассимиляционное число.
Квантовый расход – это отношение количества поглощенных квантов к количеству ассимилированных молекул СО2. Обратная величина названа квантовым выходом.
Ассимиляционное число – это соотношение между количеством СО2 и количеством хлорофилла, который содержится в листе.
Скорость (интенсивность) фотосинтеза – один из важнейших факторов, влияющих на продуктивность с/х культур, а значит и на урожай. Поэтому выяснение факторов, от которых зависит фотосинтез, должно вести к усовершенствованию агротехнических мероприятий.
Теоретически скорость фотосинтеза, как и скорость любого многостадийного биохимического процесса, должна лимитироваться скоростью самой медленной реакции. Так, например, для темновых реакций фотосинтеза нужны НАДФН и АТФ, поэтому темновые реакции зависят от световых реакций. При слабой освещенности скорость образования этих веществ слишком мала, чтобы обеспечить максимальную скорость темновых реакций, поэтому свет будет лимитирующим фактором.
Принцип лимитирующих факторов можно сформулировать следующим образом: при одновременном влиянии нескольких факторов скорость химического процесса лимитируется тем фактором, который ближе всех к минимальному уровню (изменение именно этого фактора будет непосредственно влиять на данный процесс).
Этот принцип впервые был установлен Ф. Блекманом в 1915 г. С тех пор было неоднократно показано, что разные факторы, например концентрация СО2 и освещенность, могут взаимодействовать между собой и лимитировать процесс, хотя часто один из них все же главенствует. Освещенность, концентрация СО2 и температура – вот те главные внешние факторы, влияющие на скорость фотосинтеза. Однако большое значение имеет также водный режим, минеральное питание и др.
Свет. При оценке действия света на тот или иной процесс важно различать влияние его интенсивности, качества (спектрального состава) и времени экспозиции на свету.
При низкой освещенности скорость фотосинтеза пропорциональна интенсивности света. Постепенно лимитирующими становятся другие факторы, и увеличение скорости замедляется. В ясный летний день освещенность составляет примерно 100 000 лк, а для светового насыщения фотосинтеза хватает 10 000 лк. Поэтому свет обычно может быть важным лимитирующим фактором в условиях затенения. При очень большой интенсивности света иногда начинается обесцвечивание хлорофилла, и это замедляет фотосинтез; однако в природе, растения находящиеся в таких условиях, обычно тем или иным образом защищены от этого (толстая кутикула, опущенные листья и т. п.).
Зависимость интенсивности фотосинтеза от освещенности описывается кривой, которая получила название световой кривой фотосинтеза (рис. 2.26).
|
Рис. 2.26. Зависимость интенсивности фотосинтеза от освещенности (световая кривая фотосинтеза): 1 – скорость выделения СО2 в темноте (скорость дыхания); 2 – компенсационная точка фотосинтеза; 3 – положение светового насыщения |

При слабом освещении в процессе дыхания выделяется больше СО2, чем связывается его в процессе фотосинтеза, поэтому начало световой кривой с осью абсцисс – компенсационная точка фотосинтеза, которая показывает, что в этом случае при фотосинтезе используется ровно столько СО2, сколько его выделяется при дыхании. Иными словами, со временем наступает такой момент, когда фотосинтез и дыхание будут точно уравновешивать друг друга, так что видимый обмен кислорода и СО2 прекратиться. Световая точка компенсации – это такая интенсивность света, при которой суммарный газообмен равен нулю.
Световые кривые одинаковы не для всех растений. У растений, которые растут на открытых солнечных местах, поглощение СО2 увеличивается до тех пор, пока интенсивность света не будет равна полному солнечному освещению. У растений, которые растут на затененных местах (например, кислица), поглощение СО2 увеличивается только при малой интенсивности света.
Все растения по отношению к интенсивности света делят на световые и теневые, или светолюбивые и теневыносливые. Большинство с/х растений является светолюбивыми.
У теневыносливых растений, во-первых, световое насыщение происходит при более слабом освещении, во-вторых, в них компенсационная точка фотосинтеза наступает раньше, т. е. при меньшей освещенности (рис. 2.27).
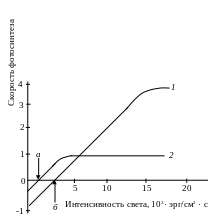
Рис.
2.27 Световые
кривые светолюбивого (1)
и теневыносливого (2)
растения (а,
б –
компенсационные точки)
Последнее связано с тем, что теневыносливые растения отличаются малой интенсивностью дыхания. В условиях слабой освещенности интенсивность фотосинтеза выше у теневыносливых растений, а при сильном свете, наоборот, – у светолюбивых.
Интенсивность света влияет и на химический состав конечных продуктов фотосинтеза. Чем выше освещенность, тем больше образуется углеводов; при низкой освещенности – больше органических кислот.
Опыты в лабораторных условиях показали, что на качество продуктов фотосинтеза влияет и резкий переход «темнота – свет» и наоборот. Сначала после включения света высокой интенсивности преимущественно образуются неуглеводные продукты из-за недостатка НАДФН и АТФ, и только через некоторое время начинают образовываться углеводы. И наоборот, после выключения света листья не сразу теряют способность к фотосинтезу, потому что на протяжении нескольких минут в клетках остается запас АТФ и НАДФ.
После выключения света сначала тормозится синтез углеводов и только потом органических веществ и аминокислот. Основная причина этого явления обусловлена тем, что торможение превращения ФГК в ФГА (и через него в углеводы) происходит раньше, чем торможение ФГК в ФЕП (и через него в аланин, малат и аспарат).
На соотношение образующих продуктов фотосинтеза влияет и спектральный состав света. Под влиянием синего света в растениях увеличивается синтез малата, аспартата и других аминокислот и белков. Эта реакция на синий свет выявлена и в С3-растениях и в С4-растениях.
С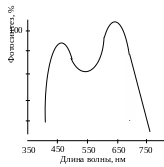 пектральный
состав света влияет и на интенсивность
фотосинтеза (рис. 2.28).
пектральный
состав света влияет и на интенсивность
фотосинтеза (рис. 2.28).
Рис.
2.28. Спектр
действия фотосинтеза
листьев
пшеницы
Спектр действия – это зависимость эффективности химического (биологического) действия света от длины его волны. Интенсивность фотосинтеза в разных участках спектра неодинакова. Максимальная интенсивность наблюдается при освещении растений теми лучами, которые максимально поглощаются хлорофиллами и другими пигментами. Интенсивность фотосинтеза наиболее высокая в красных лучах, потому что она пропорциональна не количеству энергии, а количеству квантов.
Из суммарного уравнения фотосинтеза:
6СО2 + 6Н2О → С6Н12О6 + 6О2
следует, что для образования 1 моля глюкозы нужно 686 ккал; это значит, что для ассимиляции 1 моля СО2 нужно 686 : 6 = 114 ккал. Запас энергии 1 кванта красного света (700 нм) равен 41 ккал/энштейн, а синего (400 нм) 65 ккал/энштейн. Минимальный квантовый расход при освещении красным светом равен 114 : 41 ≈ 3, а в действительности тратиться 8–10 квантов. Таким образом, эффективность использования красного света 114/41 · 8 = 34 %, а синего 114/65 ·8 = 22 %.
Концентрация СО2. Для темновых реакций нужна двуокись углерода, которая включается в органические соединения. В обычных полевых условиях именно СО2 является главным лимитирующим фактором. Концентрация СО2 в атмосфере составляет 0,045 %, но если повышать ее, то можно увеличить и скорость фотосинтеза. При кратковременном действии оптимальная концентрация СО2 составляет 0,5 %, однако при длительном воздействии возможно повреждение растений, поэтому оптимум концентрации в этом случае ниже – около 0,1 %. Уже сейчас некоторые тепличные культуры, например томаты, стали выращивать в атмосфере, обогащенной СО2.
В настоящее время большой интерес вызывает группа растений, которые намного эффективнее поглощают СО2 из атмосферы и поэтому дают более высокий урожай – так называемые С4-растения.
В искусственных условиях зависимость фотосинтеза от концентрации СО2 описывается в углекислотной кривой, которая напоминает световую кривую фотосинтеза (рис.2.29).
При концентрации СО2 0,01 % скорость фотосинтеза равна скорости дыхания (компенсационная точка). Углекислотное насыщение наступает при 0,2–0,3 % СО2, а у некоторых растениях даже при этих концентрациях наблюдается небольшое увеличение фотосинтеза.
|
Рис. 2.29. Зависимость интенсивности фотосинтеза хвои сосны от концентрации СО2 в воздухе |
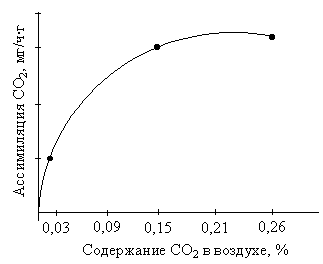
В природных условиях зависимость фотосинтеза от концентрации СО2 описывается только линейной частью кривой. Отсюда следует, что обеспеченность растений СО2 в природных условиях является фактором, который лимитирует урожай. Поэтому целесообразно выращивать растения в закрытых помещениях с повышенным содержанием СО2.
Температура оказывает заметное влияние на процесс фотосинтеза, поскольку темновые, а отчасти и световые реакции фотосинтеза контролируются ферментами. Оптимальная температура для растений умеренного климата обычно составляет около 25 оС.
Поглощение и восстановление СО2 у всех растений с повышением температуры увеличиваются, пока не будет достигнут некоторый оптимальный уровень. У большинства растений умеренной зоны снижение интенсивности фотосинтеза начинается уже после 30 оС, у некоторых южных видов после 40 оС. При большой жаре (50–60 оС), когда начинается инактивация ферментов, а также нарушается согласованность разных реакций, фотосинтез быстро прекращается. По мере повышения температуры интенсивность дыхания повышается значительно быстрей, чем интенсивность естественного фотосинтеза. Это влияет на величину наблюдаемого фотосинтеза. Зависимость интенсивности наблюдаемого фотосинтеза от температуры описывается температурной кривой, в которой выделяют три основные точки: минимум, оптимум и максимум.
Минимум – та температура при которой фотосинтез начинается, оптимум – температура, при которой фотосинтез наиболее устойчивый и достигает наибольшей скорости, максимум – та температура, после достижения которой фотосинтез прекращается (рис. 2.30).
|
Рис. 2.30. Зависимость интенсивности фотосинтеза от температуры листа: 1 – хлопчатник; 2 – подсолнечник; 3 – сорго |

Влияние кислорода. Более полувека назад было отмечено на первый взгляд парадоксальное явление. Кислород воздуха, который является продуктом фотосинтеза, является одновременно и его ингибитором: выделение кислорода и поглощение СО2 падают по мере увеличения концентрации О2 в воздухе. Этот феномен назвали именем его открывателя – эффект Варбурга. Этот эффект присущ всем С3-растениям. И только в листьях С4-растений его не удалось выявить. Сейчас твердо установлено, что природа эффекта Варбурга связана с оксигеназными свойствами основного фермента цикла Кальвина – РДФ-карбоксилазы. При большой концентрации кислорода начинается фотодыхание. Установлено, что при снижении концентрации О2 до 2–3 % фосфогликолат не образуется, исчезает и эффект Варбурга. Таким образом, оба эти явления – проявление оксигеназных свойств РДФ-карбоксилазы и образование гликолата, а также уменьшение фотосинтеза в присутствии О2 тесно связаны один с другим.
Очень низкое содержание О2 или полное отсутствие, как и увеличение концентрации до 25–30 %, тормозит фотосинтез. Для большинства растений некоторое снижение природной концентрации (21 %) О2 активирует фотосинтез.
Влияние оводненности тканей. Как уже отмечалось, вода участвует в световой стадии фотосинтеза как донор водорода для восстановления СО2. Однако, роль лимитирующего фотосинтез фактора играет не минимальное количество воды (приблизительно 1 % поступившей), а та вода, которая входит в состав клеточных мембран и является средой для всех биохимических реакций, активирует ферменты темновой фазы. Кроме того, от количества воды в замыкающих клетках зависит степень открытия устьиц, а тургорное состояние всего растения определяет расположение листьев по отношению к солнечным лучам. Количество воды косвенно влияет на изменение скорости отложения крахмала в строме хлоропласта и даже на изменение структуры и расположение тилакоидов в строме.
Зависимость интенсивности фотосинтеза от оводненности тканей растений, как и зависимость от температуры, описывается переходной кривой, имеющей три основные точки: минимум, оптимум и максимум.
При обезвоживании меняется не только интенсивность фотосинтеза, но и качественный состав продуктов фотосинтеза: меньше синтезируется малата, сахарозы, органических кислот; больше – глюкозы, фруктозы аланина и других аминокислот.
К тому же установлено, что при нехватке воды в листьях накапливается АБК – ингибитор роста.
Концентрация хлорофилла, как правило, не бывает лимитирующим фактором, однако количество хлорофилла может уменьшаться при различных заболеваниях (мучнистая роса, ржавчина, вирусные болезни), недостатке минеральных веществ и с возрастом (при нормальном старении). Когда листья желтеют, говорят, что они становятся хлоротичными, а само явление называют хлорозом. Хлоротические пятна на листьях часто бывают симптомом заболевания или недостатка минеральных веществ.
Хлороз может быть вызван и недостатком света, так как свет нужен для конечной стадии биосинтеза хлорофилла.
Минеральные элементы. Для синтеза хлорофилла нужны и минеральные элементы: железо, магний и азот (два последних элемента входят в его структуру), потому они особенно важны для фотосинтеза. Важен также калий.
Для обычного функционирования фотосинтетического аппарата растение должно быть обеспечено необходимым количеством (оптимальным) минеральных элементов. Магний, кроме того, что входит в состав хлорофилла, участвует в действии сопрягающих белков при синтезе АТФ, влияет на активность реакций карбоксилирования и восстановление НАДФ+.
Железо в восстановленной форме необходимо для процессов биосинтеза хлорофилла и железосодержащих соединений хлоропластов (цитохромов, ферредоксина). Нехватка железа нарушает циклическое и нециклическое фотофосфорилирование, синтез пигментов, изменение структуры хлоропластов.
Марганец и хлор принимают участие в фотоокислении воды.
Медь входит в состав пластоцианина.
Недостаток азота оказывает влияние не только на формирование пигментных систем и структур хлоропластов, но и на количество и активность РДФ-карбоксилазы.
При недостатке фосфора нарушаются фотохимические и темновые реакции фотосинтеза.
Калий играет полифункциональную роль в ионной регуляции фотосинтеза, при его недостатке в хлоропластах разрушается структура гран, устьица слабо открываются на свету и недостаточно закрываются в темноте, ухудшается водный режим листа, т. е. нарушаются все процессы фотосинтеза.
Возраст растений. Только после создания фитотронов, где можно выращивать растения в контролируемых условиях, удалось получить надежные результаты. Выявлено, что во всех растениях только в самом начале жизненного цикла, когда формируется фотосинтетический аппарат, интенсивность фотосинтеза увеличивается, очень быстро достигает максимума, затем немного уменьшается и дальше меняется очень мало. Например, у злаков фотосинтез достигает максимальной интенсивности в фазу кущения. Это объясняется тем, что максимальная фотосинтетическая активность листа совпадает с окончанием периода его формирования. Затем начинается старение и уменьшение фотосинтеза.
Интенсивность фотосинтеза зависит в первую очередь от структуры хлоропластов. При старении хлоропластов разрушаются тилакоиды. Доказывают это с помощью реакции Хила. Она идет тем хуже, чем больший возраст хлоропластов. Таким образом, показано, что интенсивность определяется не количеством хлорофилла, а структурой хлоропласта.
В оптимальных условиях влажности и азотного питания снижение фотосинтеза с возрастом происходит медленнее, так как в этих условиях хлоропласты медленнее стареют.
Генетические факторы. Процессы фотосинтеза в определенной степени зависят от наследственности растительного организма. Интенсивность фотосинтеза различна у растений разных систематических групп и жизненных форм. У трав интенсивность фотосинтеза выше, чем у древесных растений (табл. 2.5).
Таблица 2.5