

2.10. Фотодыхание
Путь восстановления СО2 при фотосинтезе М. Кальвин изучал при относительно высоких концентрациях углекислого газа. При более низких концентрациях СО2, приближающихся к атмосферным (0,045 %), в хлоропласте образуется значительное количество фосфогликолевой кислоты. Это связано с тем, что в условиях низкого парциального давления СО2 и высокого парциального давления О2 РДФ-карбоксилаза может присоединять к рибулозо-1,5-дифосфату кислород; это значит, что она может выполнять не только карбоксилирующую, но и оксигенерирующую функции. Присоединение кислорода к молекуле рибулозодифосфата приводит к тому, что вместо двух молекул ФГК образуется одна молекула ФГК и одна молекула фосфогликолата. Последняя во время транспорта через мембрану хлоропласта дефосфорилируется при участии фосфогликолатфосфатазы и превращается в гликолат, который поступает в специальную органеллу-периксисому (рис. 2.25).
В последней содержится каталаза – фермент, разлагающий перекиси. В периксисоме гликолат вступает в реакцию с кислородом воздуха, в результате образуется глиоксилат и перекись водорода. Реакцию катализирует гликолатоксидаза.
Перекись сразу разрушается каталазой на воду и кислород, а глиоксилат в результате реакции трансаминирования, которая проходит при участии глиоксилатглицинаминотрансферазы, превращается в глицин, который перемещается в митохондрию. Тут из двух молекул глицина при участии серинглиоксилатаминотрансфераза синтезируется одна молекула серина, образуется СО2, аммиак и восстанавливается НАД+. СО2 или выделяется в атмосферу, это значит теряется для растения, или вновь может связаться в С3-цикле.
|
Рис. 2.25. Реакции фотодыхания |
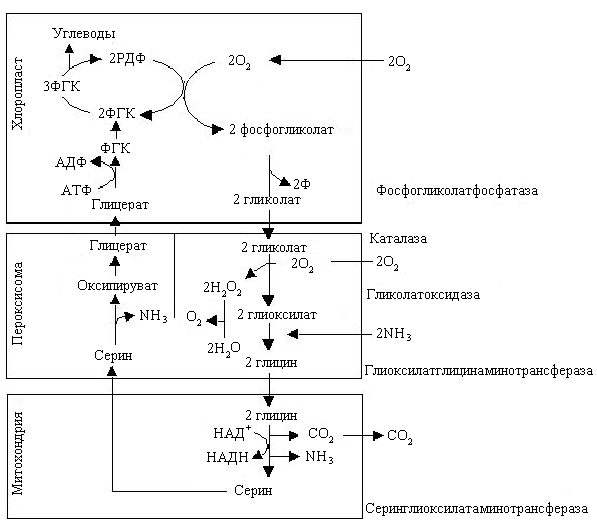
Предполагают, что образующийся НАДН может быть связан с электрон-транспортной цепью дыхания, что приводит к синтезу АТФ при его окислении.
Серин из митохондрий транспортируется вновь в периксисому, где после дезаминирования превращается в оксипируват, потом в глицерат, и наконец, в ФГК. ФГК – конечный продукт этого цикла может поступать в С3-цикл. Цикл превращения гликолата в триозофосфат называется гликолатным. Образовавшийся аммиак не выделяется в окружающую среду, а связывается в виде глутамина.
Так как образуются кислоты, содержащие 2 атома углерода, то этот цикл получил название С2-цикла. С другой стороны, так как во время этих превращений происходит поглощение кислорода и выделение СО2, то этот процесс получил название фотодыхания.
Фотодыхание (или световое дыхание) – это индуцированное светом поглощение О2 и выделение СО2, которое наблюдается в зеленых клетках. Интенсивность фотодыхания увеличивается при повышении освещенности.
Хотя органеллой фотодыхания считается периксисома, в этом цикле непосредственное участие принимают хлоропласты и митохондрии. В клетке эти органеллы примыкают друг к другу, что также подчеркивает их функциональную зависимость. Таким образом, мы приходим к очень важному выводу: в клетке вместе с изолированностью органелл существует их интеграция.
Не у всех растений фотодыхание в равной степени идет интенсивно. У сои оно идет с большей скоростью, а у сахарного тростника его долгое время не могли выявить. Вообще, у С3-растений фотодыхание более интенсивнее, чем в С4-растениях.
Интенсивное фотодыхание может снизить продуктивность С3-растений на 30 %, так как при фотодыхании происходит не поглощение СО2, а его потери. Из каждых 10 молекул риболозо-1,5-дифосфата, образовавшегося в С3-цикле и включившихся в фотодыхание, одна разрушается:

В связи с этим важной задачей для физиологов и селекционеров растений считается выведение сортов с/х растений, у которых в максимальной степени проявляется карбоксилазная и подавлена оксигеназная реакция, и таким образом, подавлено фотодыхание.
Низкую интенсивность фотодыхания в С4-растениях объясняют тем, что в них высокая концентрация СО2 в клетках-обкладки. В этих растениях РДФ-карбоксилаза находится только в клетках-обкладки, где и происходит процесс. Некоторые исследователи (Мокроносов, 1984 г.) считают, что в С4-растениях фотодыхание подавлено из-за недостатка О2 в клетках обкладках. В этих клетках не идет фотолиз воды, так как они не имеют ФС II, а поступление атмосферного кислорода затрудненно, потому что эти клетки расположены в глубине листовой пластинки и между ними почти нет межклетников.
В настоящее время фотодыхание рассматривают как одну из необходимых функций зеленого растения. Во-первых, фотодыхание, как и С3-цикл, САМ-цикл и другие процессы, является источником метаболитов для разных синтезов. Во-вторых, почти доказано, что при образовании серина и глицина восстанавливается митохондриальная НАД+. В-третьих, выявлена связь фотодыхания с ассимиляцией азота. И, наконец, появилась теория, в соответствии с которой излишки продуктов цикла Кальвина служат сигналом, который увеличивает фотосинтез. Таким образом, соотношение фотосинтеза и фотодыхания выполняют регуляторную роль в растениях.