

2.9. Продукты темновой стадии фотосинтеза
Образовавшиеся в процессе темновой стадии вещества делят на первичные, промежуточные и конечные. Раньше считали, что первичным продуктом фотосинтеза является крахмал. Затем, в 40-е годы прошлого века шла дискуссия, где одни ученые в качестве первичных продуктов фотосинтеза считали сахарозу, другие – фруктозу и глюкозу. В 50-е годы, когда стали использовать изотопы (радиоактивный СО2), то показали, что через несколько секунд после попадания света на лист или зеленую клетку водоросли метка у С3-растений появляется в ФГК, а затем в продуктах ее восстановления – фосфорилированных сахарах, составных компонентах цикла Кальвина. С того времени ФГК стала считаться первичным продуктом ассимиляции СО2.
Дальнейший путь превращения первичных продуктов фотосинтеза может быть разным в зависимости от вида растений, их физиологического состояния, условий питания, температуры, освещенности. Все это приводит к образованию конечных продуктов разного состава и качества. Некоторые из этих конечных продуктов (крахмал) образуется в самих хлоропластах.
Крахмал используется для синтеза разных веществ, в том числе сахарозы – главного по количеству сахара, запасаемого в растениях. Сахароза синтезируется в слое цитоплазмы, которая прилагает к хлоропласту.
Качественный состав конечных продуктов фотосинтеза зависит от спектрального состава света. Опыты с выращиванием растений при освещении только красным или только синим светом, показали, что при действии красного света увеличивается синтез крахмала; при действии синего – уменьшается. При облучении синим светом увеличивается проницаемость мембраны хлоропласта для ФГК и неорганического фосфата: ФГК выходит, а неорганический фосфат входит в хлоропласт. В результате соотношение ФГК/Фн в хлоропласте уменьшается. Это подавляет синтез крахмала. ФГК, которая вышла из хлоропласта, используется на синтез амино- и органических кислот. «Фотосинтетическое» происхождение (на основе ФГК из С3-цикла) амино- и органических кислот впервые доказано в СССР в 50-е годы А. А. Ничипоровичем и др. Минеральные элементы также влияют на качественный и количественный состав синтезируемых веществ. Например, у кукурузы (С4-растение) в условиях аммонийного питания образование малата понижается, а аспартата увеличивается.
При фотосинтезе, как мы отмечали, действуют две цепи переноса электронов: циклическое фотофосфорилирование (ФС I) и нециклическое (ФС I и ФС II). Обе системы поставляют АТФ, а ФС II еще и восстановительную силу (НАДФН) для фиксации СО2 и восстановления его до уровня углеводов. В других клеточных реакциях используется АТФ, который образуется главным образом в процессе окислительного фотофосфорилирования.
Окислительное фосфорилирование связано с процессом дыхания и осуществляется при помощи другой цепи переноса электронов, которую мы рассмотрим позже.
Постепенно, т. е. в несколько этапов, должно осуществляться не только разрушение органических молекул (так как это необходимое условие эффективного запасания и использования энергии), но и синтез сложных соединений – белков, нуклеиновых кислот, липидов или полисахаридов. Когда из простых молекул строятся более крупные, этим простым молекулам часто необходима активация; другими словами, им необходима энергия. Поэтому синтез того или иного соединения проходит более сложным путем, чем его распад.
Синтез сахарозы и полисахаридов довольно сложный процесс. Углерод, фиксированный при фотосинтезе, как уже отмечалось, превращается в фотосинтезирующих клетках в два основных углевода: сахарозу и крахмал.
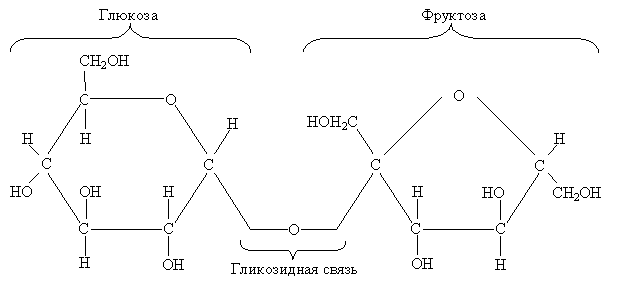
Сахароза. Большая часть углеводов выносится из клетки, вероятно, в виде сахарозы. Сахара транспортируются по растению в виде дисахарида, который состоит из остатков глюкозы и фруктозы.
Сахароза транспортируется в те части растения, где она особенно нужна – к участкам быстрого роста, развивающимся семенам и клубням.
|
Коротко путь синтеза сахарозы у фотосинтезирующих клеток С3-растений следующий. Фруктоза-6-фосфат под влиянием специфических ферментов превращается в глюкоза-6-фосфат и глюкоза-1-фосфат, причем легко происходит и обратное превращение;
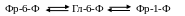
Из этих трех гексозофосфатов строятся затем цепи углеводных молекул, которые используются для транспорта, хранения и в других реакциях синтеза. Для таких превращений гексозофосфаты должны быть активированы. Это обычно достигается их присоединением к нуклеотидам – сложным кольцевым структурам, подобным адениловой кислоте.
Продуктом такой реакции присоединения являются нуклеотидные производные моносахаридов или нуклеотидсахаров. Чаще других встречается уридиндифосфатглюкоза (УДФГл), которая образуется в реакциях между уридинтрифосфатом (УТФ) и глюкоза-1-фосфатом (Гл-1-Ф). Сам УТФ образуется косвенным путем, в результате переноса фосфатной группы от АТФ на УДФ:
УДФ + АТФ → УТФ + АДФ
У
пирофосфат
Нуклеотиды АТФ и УТФ присутствуют во всех клетках, потому что они используются вместе с другими нуклеотидами в синтезе ДНК и РНК. Сахароза (ГлФр) образуется в реакции между УДФГл и Фр-6-Ф:
|
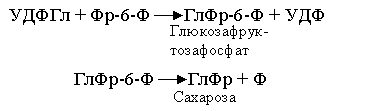
Равновесие этой реакции сильно сдвинуто в сторону синтеза сахарозы, что обеспечивает запасание данного дисахарида в значительных концентрациях. Для последующего использования сахароза должна сначала подвергнуться расщеплению (фермент инвертаза катализирует ее гидролиз с образованием свободной глюкозы и фруктозы):
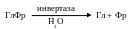
Энергия гликозидной связи в такой реакции растрачивается впустую, распределяясь между двумя молекулами. Поэтому, когда глюкозе и фруктозе предстоит распад в процессе дыхания или участие (в качестве сырья) в синтезе полисахаридов, то они должны сначала снова подвергнуться фосфорилированию за счет АТФ.
Крахмал представляет собой как бы временный запас фиксированного углевода. Он образуется во время наиболее активного периода фотосинтеза.
Длинные полимерные цепи крахмала построены из элементарных звеньев – остатков глюкозы:
|
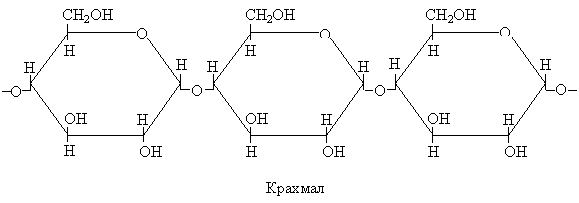
Крахмал представлен двумя формами: линейным полимером – амилазой, и разветвленным полимером – амилопектином. Крахмал – основной запасной полисахарид растений. Он нерастворим в воде и откладывается слой за слоем в крахмальных зернах в хлоропластах, или накапливается в лейкопластах в тканях стебля, корней, семян. Он просто обнаруживается, поскольку способен легко окрашиваться йодом в синий цвет. Так как крахмал нерастворим в воде в отличие от сахарозы и гексоз, он не вызывает в клетках осмотического эффекта. Поэтому образование крахмала в клетках листа в периоды интенсивного фотосинтеза, не вызывает его подавления, которое происходит в результате накопления других продуктов фотосинтеза (ассимилятов). В темноте крахмал постепенно снова гидролизуется с образованием глюкозофосфата, который затем превращается в сахарозу, которая транспортируется в другие части растений.
Начальным продуктом для синтеза крахмала служит аденозиндифосфатглюкоза (АДФГл), образующаяся из АТФ и Гл-1-Ф. Молекула крахмала строится путем поэтапного присоединения одного глюкозного остатка к другому в реакции АДФГл с преобразованной глюкозной цепью:
А
крахмал
При низком содержании глюкозы крахмал разрушается и переводится в сахарозу. Однако сначала он разрушается до остатков глюкозы и к каждому из них присоединяется остаток фосфорной кислоты, т. е. образуется Г-1-Ф, чем и обеспечивается сохранение энергии связи:
[Глюкозаn+1] + Ф → [Глюкозаn] + Гл-1-Ф.
Затем Гл-1-Ф может использоваться для синтеза сахарозы. В семенах и некоторых других органах, в которых одновременно идет разрушение больших количеств крахмала, он распадается до дисахарида мальтозы под воздействием α-амилазы. Мальтоза затем разрушается до глюкозы:
|

Из последней может снова синтезироваться сахароза. Для превращения глюкозы в глюкоза-6-фосфат необходима АТФ.
Целлюлоза служит основным компонентом первичной клеточной оболочки и является одним их наиболее распространенных углеводов на Земле. Молекулы целлюлозы построены подобно молекулам крахмала из остатков глюкозы, только в отличие от последнего соединены по другому:
|
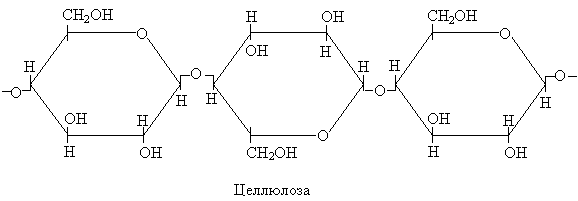
Эти структурные отличия обусловлены тем, что два рассматриваемых полимера глюкозы (глюканы) значительно отличаются по своей природе; крахмал, например, легко переваривается в организме человека, а целлюлоза совсем не переваривается. Основная разница между крахмалом и целлюлозой состоит в том, что 1-й и 4-й углеродные атомы двух соседних остатков глюкозы соединены в крахмале α-связями, а целлюлозе – β-связями.
Отличается от крахмала и реакция синтеза целлюлозы. Роль донора глюкозы выполняет нуклеотидное производное моносахарида – гуаназиндифосфатглюкоза (ГДФГ) и связь между мономерными звеньями относится к β-типу:
ГнДА + АТФ → ГнТФ + АДФ
ГнТФ + Г-1-Ф → ГнДФГ + ФФн
ГнДФГ + [Глюкозаn] → ГнДФ + [Глюкозаn+1]
целлюлоза
В некоторых случаях донором глюкозы для синтеза целлюлозы может быть и уридиндифосфатглюкоза.
У высших растений целлюлоза разрушается редко (если не считать разрушений под действием микробов). Два известных исключения из правил касаются клеток в отделительной зоне листа, образованной перед сбрасыванием листьев, и поперечных стенок сосудов ксилемы. В отделительной зоне листа фермент целлюлаза разрушает клеточные стенки, расщепляет образующуюся в них целлюлозу до отдельных мономерных единиц, т. е. до глюкозы. Клеточные оболочки, в конце концов, разрываются, и лист сбрасывается.
Целлюлозные микрофибриллы в клеточной оболочке скреплены при помощи матрикса из смешанных полисахаридных цепей (пентозы). Эти полисахариды синтезируются также из предшественников, нуклеотидсахаридов, главным образом, в диктиосомах. Пузырьки, которые отшнуровываются от диктиосом, в конце концов, сливаются с плазмалеммой и таким образом передают свое содержимое образующейся клеточной оболочке.
Все полисахариды легко переходят один в другой, но синтез их всегда идет через стадию нуклеотидсахаров, тогда как разрушение проходит более прямым путем.