

2.8. Темновая фаза
Темновая фаза фотосинтеза – это комплекс ферментативных реакций, во время которой происходит восстановление поглощенного углекислого газа за счет продуктов световой фазы (АТФ и НАДФН). Различают несколько циклов восстановления СО2.
Цикл Кальвина. Этот способ ассимиляции СО2 является основным и присущ всем растениям. Он был расшифрован американскими учеными во главе с М. Кальвином. В 1961 году М. Кальвину за установление последовательности реакций в этом цикле и была присуждена Нобелевская премия.
Этот цикл начинается с присоединения СО2 к акцептору – пятиуглеродному сахару рибулозо-1,5-дифосфату (РДФ). Присоединение СО2 к тому или ионному веществу называется карбоксилированием, а фермент катализирующий такую реакцию – карбоксилазой.
В данной реакции карбоксилирование происходит с участием фермента рибулозодифосфаткарбоксилазы (РДФ-карбоксилаза). Это самый распространенный в мире фермент.
Продукт реакции, содержащий 6 атомов углерода, в присутствии воды сразу распадается на две молекулы 3-фосфоглицириновой кислоты (3-ФГК):

С данной реакции и начинается цикл Кальвина. ФГК и является, по современным взглядам, первичным продуктом ассимиляции углерода.
Для дальнейших превращений ФГК необходимы вещества световой фазы фотосинтеза: АТФ и НАДФН. Сначала 3-ФГК фосфорилируется при участии АТФ и образуется 1,3-дифосфоглицириновая кислота. Реакция катализируется ферментом фосфоглицераткиназой:

Затем происходит восстановление за счет НАДФН и образуется фосфоглицириновый альдегид ФГА:

Суммарный результат второй стадии: восстановление карбоксильной группы кислоты (–СООН) до альдегидной (–СНО).
Процесс превращения катализируется дегидрогеназой фосфоглициринового альдегида. Дальнейшее превращение фосфоглициринового альдегида может происходить 4 путями.
ФГА частично с помощью триозофосфатизомеразы превращается в фосфодиоксиацетон (ФДА):

Это первый путь превращения ФГА.
Таким образом, в клетку поступают две найпростейшие формы сахаров: альдоза (ФГА) и кетоза (ФДА). Это трехуглеродные сахара (триозосахара) с присоединенной к ним фосфатной группой содержат больше химической энергии, чем ФГК. Это первые углеводы, которые образуются при фотосинтезе.
С помощью альдолазы фосфодиоксиацетон (ФДА) соединяется с другой молекулой ФГА и образуется молекула фруктозо-1,6-дифосфата (ФДФ).
|
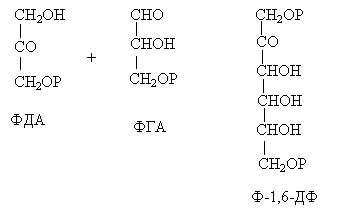

Это второй путь превращения ФГА.
Фруктозо-1,6-дифосфат дефосфорилируется и превращается во фруктозо-6-фосфат (Ф-6-Ф), что сопровождается накоплением в среде неорганического фосфата. Фруктозо-6-фосфат в дальнейшем может выйти из цикла и использоваться для синтеза запасных форм углеводов: сахарозы, крахмала, других полисахаридов.
Однако ФГА (третий путь) может реагировать с эквимолярным количеством Ф-6-Ф, в результате образуются равные количества ксилулозо-5-фосфата и эритрозо-4-фосфата (транскетолаза). Затем эритрозо-4-фосфат реагирует с равным количеством ФДА и образуется седагептулозо-1,7-дифосфат (альдолаза), которая фосфорилируется до седагептулозо-7-фосфата с участием седагептулозодифосфатазы.

Четвертый путь превращения ФГА связан с его реакцией с седагептулозо-7-фосфатом с образованием равных (эквимолярных) количеств рибозо-5-фосфата и ксилулозо-5-фосфата. Ксилулозо-5-фосфат эпимиризуется, а рибозо-5-фосфат изомерезуется до рибулозо-5-фосфата, последняя фосфорилируется за счет АТФ и образуется рибулозо-1,5-дифосфат – первичное соединение цикла Кальвина (акцептор СО2). В этих реакциях тратится еще три молекулы АТФ.
И з
приведенных реакций цикла Кальвина
видно, что фотосинтез, являясь процессом
запасания энергии, тем не менее, для
своего существования требует затраты
энергии.
з
приведенных реакций цикла Кальвина
видно, что фотосинтез, являясь процессом
запасания энергии, тем не менее, для
своего существования требует затраты
энергии.
В цикле Кальвина образование фруктозо-6-фосфата можно представить в виде следующего суммарного выражения:
6СО2 + 12НАДФН + 12Н+ + 18АТФ + 11Н2О →
фруктозо-6-фосфат + 12НАДФ+ + 18АДФ + 17Фн
18 молекул АТФ запасают около 140 ккал и 12 НАДФН – ~ 615 ккал. Следовательно, поглощено около 755 ккал энергии. При этом в гексозах запасается около 670 ккал/моль. При таком балансе КПД составляет около 90 %. ~ 10 % энергии растрачивается на поддержание цикла.
АТФ и НАДФН, которые образуются в световой стадии и используются на восстановление СО2, получили название ассимиляционной силы.
Цикл Кальвина подразделяют на три фазы:
– карбоксилирующую РДФ + СО2 → 2ФГК;
– восстановительную ФГК → ФГА;
– регинирующую ФГА → РДФ.
Каждая шестая молекула ФГА выходит из цикла и используется на синтез сахарозы или полисахаров, тогда как остальные 5 молекул через приведенные выше промежуточные реакции преобразуются в три молекулы рибулозо-1,5-дифосфата. Так как первичный продукт цикла Кальвина – ФГК – содержит три атома углерода, то этот цикл получил название С3-цикла ассимиляции СО2. Упрощенную схему цикла можно представить в виде (рис.2.18):
|
Рис.2.18. Упрощенная схема цикла Кальвина |
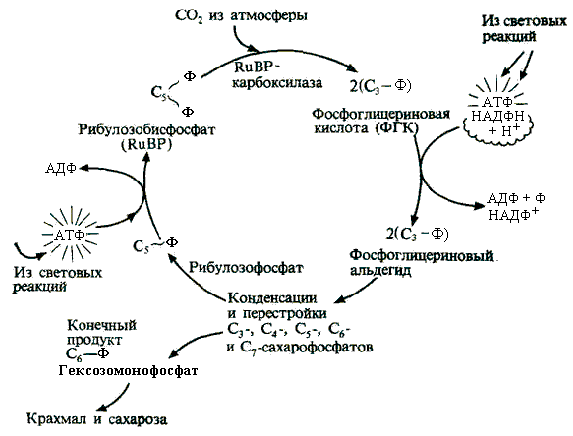
Последовательность реакций на пути преобразования СО2 в сахар удалось выявить благодаря использованию радиоактивного углерода 14С и хромотографии на бумаге.
Описанный цикл восстановления СО2 до сахаров локализован в хлоропластах, так же как и биосинтез крахмала из образованных в них гексозофосфатов. «Главный» же по количеству сахар, запасенный в растительной клетке – сахароза, – синтезируется уже вне хлоропласта: в слое цитоплазмы, прилегающем к наружной мембране этой органеллы. Сахароза синтезируется из Ф-6-Ф, образованной из ФГА и ФДА, которые в отличие от других сахаров цикла (пентоз и гексоз) легко транспортируются через мембраны хлоропластов.
Скорость цикла Кальвина зависит не только от количества образованных в световой стадии АТФ и НАДФН, но и от их соотношений. Только соотношение 3АТФ и 2НАДФН обеспечивает активное восстановление углерода и запасание энергии.
Когда степень сопряжения работы ЭТЦ фотосинтеза с фотофосфорилированием мала, тогда интенсивность фотосинтеза, в первую очередь, может снизиться за счет уменьшения количества рибулозо-1,5-дифосфата, так как в этом случае будет лимитироваться фосфорилирование рибулоза-5-фосфата.
Кроме этого, в цикле при недостатке АТФ и НАДФН уменьшается возможность восстановления ФГК до триоз и поэтому одновременно со снижением интенсивности фотосинтеза среди ассимилятов (продуктов фотосинтеза) клетки увеличивается часть неуглеродных соединений. Такое явление характерно, например, для растений, выращенных при слабом освещении.
Цикл Хетча – Слэка. Цикл Кальвина – основной, но не единственный путь восстановления СО2. Так австралийские ученые М. Хетч и К. Слэк (1966) и советский ученый Ю. Карпилов (1960) выявили, что у некоторых растений, главным образом, тропических и субтропических, таких как кукуруза, сахарный тростник, сорго и другие, основная часть меченного углерода (14СО2) уже после нескольких секунд фотосинтеза обнаруживается не в фосфоглицериновой кислоте, а в щавелево-уксусной (ЩУК), яблочной (ЯК) и аспаргиновой (АК) кислотах. В этих кислотах можно обнаружить в первые секунды до 90 % поглощенного 14СО2. Через 5–10 минут метка появлялась в фосфоглицериновой кислоте, а затем в фосфоглицериновых сахарах. Так как эти органические кислоты содержат по 4 атома углерода, то такие растения начали называть С4-растениями в отличие от С3-растений, в которых радиоуглеродная метка появляется, прежде всего, в ФГК.
Это открытие положило начало серии исследований, в результате которых подробно был изучен химизм превращения углерода в фотосинтезе в С4-растениях. Акцептором углекислого газа в этих растениях является фосфоенолпировиноградная кислота (ФЕП) (рис.2.19).
|
Рис. 2.19 С4 – путь фотосинтеза |
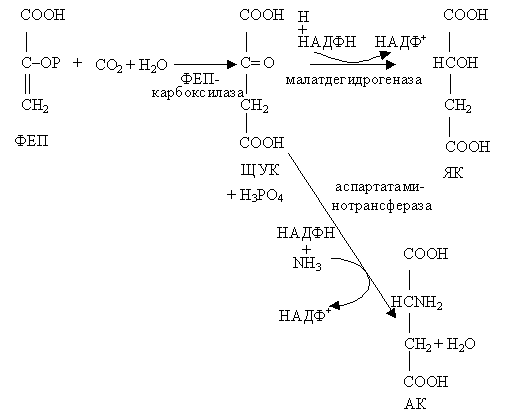
ФЕП образуется из пировиноградной или 3-фосфоглицериновой кислоты. В результате β-карбоксилирования ФЕП превращается в четырехуглеродную щавелевоуксусную кислоту:
ФЕП + СО2 + Н2О → ЩУК + Фн
Фермент, который катализирует присоединение СО2 к ФЕП – фосфоенолпируваткарбоксилаза – найден сейчас у многих одно- и двудольных растений. Образовавшаяся ЩУК при участии НАДФН (продукта световой реакции фотосинтеза) восстанавливается до яблочной кислоты (малата):
ЩУК + НАДФН + Н+ → малат + НАДФ+
Реакция катализируется НАДФ+-зависимой малатдегидрогеназой, локализованной в хлоропластах клеток мезофилла.
У некоторых растений, образовавшаяся ЩУК в процессе восстановительного аминирования с участием аспартатаминотрансферазы преобразуется в аспарагиновую кислоту. В дальнейшем яблочная (или аспаргиновая) кислоты декарбоксилируются, образуется СО2 и трехуглеродное соединение. СО2 включается в цикл Кальвина, присоединяясь к рибулозо-5-фосфату, а трехуглеродное соединение используется для регенерации фосфоенолпирувата.
В настоящее время в зависимости от того, какая из органических кислот (малат или аспартат) декарбоксилируется, С4-растения делят на два типа: малатный тип (кукуруза, тростник) и аспартатный (сорго и др.) типы.
В свою очередь растения последнего типа подразделяют на две группы: растения, которые используют в реакции НАД-зависимую малатдегидрогеназу и растения, которые используют фосфоенолпируват-карбоксикиназу.
Как отмечалось, С4-растения отличаются от С3-растений и по анатомии листовой пластины. Фотосинтез идет в клетках обкладки и в клетках мезофилла. Оба типа фотосинтезирующих тканей отличаются по строению хлоропластов. Хлоропласты клеток мезофилла имеют строение, присущее большинству растений: они содержат два типа тилакоидов – тилакоиды гран и тилакоиды стромы (гранальные хлоропласты). Клетки обкладки содержат более крупные хлоропласты, часто наполненные крахмальными зернами и не имеющими гран, т. е. эти хлоропласты содержат только тилакоиды стромы (агранальные).
Считают, что агранальные хлоропласты образуются в процессе онтогенеза листа из обычных гранальных хлоропластов, так как на ранних стадиях развития эти хлоропласты также имеют граны.
Таким образом, для С4-растений характерны следующие особенности в строении:
– многочисленные воздушные полости, по которым воздух из атмосферы подходит непосредственно к большому количеству фотосинтезирующих клеток, обеспечивая эффективное поглощение углекислоты;
– слой клеток обкладки сосудистых пучков, плотно упакованных около проводящих пучков;
– клетки мезофилла, которые располагаются менее плотными слоями около клеток обкладки сосудистых пучков;
– большое количество плазмодесм между клетками обкладки сосудистых пучков и клетками мезофилла;
– содержание как гранальных, так и агранальных хлоропластов.
Для малатного типа растений характерен следующий путь фотосинтеза. Гранальные и агранальные хлоропласты отличаются и по характеру проходящих в них реакций фотосинтеза. В клетках мезофилла с мелкими гранальными хлоропластами происходит карбоксилирование ФЕП с образованием ЩУК (первичное карбоксилирование), а затем образуется малат. Малат перемещается в клетки обкладки. Тут малат окисляется и декарбоксилируется с участием малатдегидрогеназы. Образуется СО2 и пируват. СО2 используется для карбоксилирования рибулозо-1,5-дифосфата (вторичное карбоксилирование) и таким образом включается С3-цикл, который идет в агранальных хлоропластах клеток обкладки. Пируват возвращается в клетки мезофилла, где фосфорилируется за счет АТФ, что приводит к регенерации ФЕП, и цикл замыкается (рис. 2.20).
|
Рис. 2.20. Фотосинтез С4-растений, которые используют в реакциях декарбоксилирования НАДФ-зависимый «яблочный фермент» (малатдегидрогеназа*) |

Таким образом, в С4-растениях карбоксилирование происходит два раза: в клетках мезофилла и в клетках обкладки.
Что происходит в растениях, в которых из ЩУК образуется аспартат (аспарагиновая кислота)? У тех растений, которые используют для реакций декарбоксилирования НАД-зависимую малатдегидрогеназу, ЩУК трансаминируется под воздействием цитоплазматической аспартаминотрансферазы, которая использует в качестве донора аминогруппы глутаминовой кислоты (ГК-глутаминовая, ОГК-2-оксоглутаминовая кислота) (рис. 2.21).
|
Рис. 2.21. Фотосинтез С4-растений, которые используют в реакциях декарбоксилирования НАД-зависимый «яблочный фермент» (малатдегидрогеназа):
|
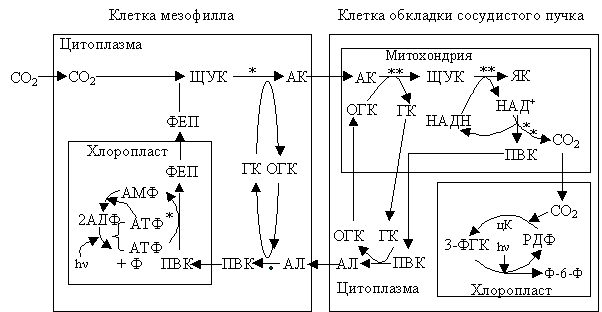

Образовавшаяся АК переходит из цитоплазмы клеток мезофилла в митохондрии клеток обкладки сосудистого пучка, вероятно, через плазмодесмы. Там происходит противоположная реакция трансаминирования, которая приводит к образованию ЩУК. Затем митохондриальная малатдегидрогеназа восстанавливает ЩУК до ЯК. ЯК декарбоксилируется НАД-зависимой малатдегидрогеназой с образованием ПВК и СО2.
СО2 диффундирует из митохондрий в хлоропласты, где включается в цикл Кальвина. ПВК поступает в цитоплазму, где трансаминируется аминотрансферазой и превращается в аланин (АЛ); донором аминогруппы является ГК.
Аланин переносится из цитоплазмы клеток обкладки в цитоплазму клеток мезофилла (вероятно, через плазмодесмы). В дальнейшем он преобразуется в ПВК (аланинаминотрансфераза); акцептором аминогруппы является ОГК (оксаглутаровая кислота). Затем ПВК переходит в хлоропласты мезофилла и превращается в ФЕП.
В С4-растениях, использующих в реакции декарбоксилирование фосфоенолпируват-карбоксикиназу, последовательность реакций напоминает предыдущие. Только в этом случае ЩУК декарбоксилируется фосфоенолпируваткарбоксикиназой с образованием СО2 и ФЕП (рис. 2.22).
|
Рис. 2.22. Фотосинтез С4-растений, использующих в реакции декарбоксилирования фермент фосфоенолпируват-карбрксикиназу* |
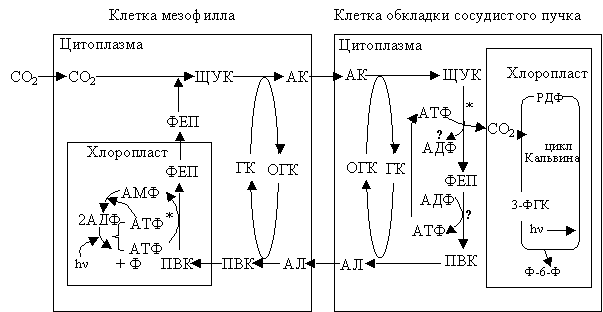
Внутриклеточная локализация ФЕП-карбоксикиназы и аспартат-аминотрансферазы в этом случае пока неизвестна. Неизвестна и судьба ФЕП; однако, как считают, он превращается в ПВК (эти реакции обозначены вопросительным знаком).
Образовавшийся в ходе реакции СО2 используется в качестве субстрата в реакциях цикла Кальвина в хлоропластах клеток обкладки сосудистого пучка. Аланин из клеток обкладки пучка превращается в ФЕП в клетках мезофилла листа; это происходит в той же последовательности реакций, что и в предыдущих циклах.
В настоящее время пришли к выводу, что основная функция С4-цикла, который идет в клетках мезофилла листа – концентрирование СО2 для С3-цикла. С4-цикл является своеобразным насосом – «углекислотной помпой». Находящаяся в мезофильных клетках ФЕП-карбоксилаза очень активна. Она может фиксировать СО2, включая его в органические кислоты при более низких концентрациях СО2, чем это делает РДФ-карбоксилаза, и активность последней у С4-растений небольшая. Благодаря функционированию этой углекислотной помпы у С4-растений концентрация СО2 в клетках обкладки, где идет цикл Кальвина, в несколько раз выше, чем в среде. Это очень важно, так как С4-растения живут в условиях повышенных температур, когда растворимость СО2 значительно ниже.
Кооперация между двумя циклами связана не только с перекачкой СО2. Для восстановления ФГК в цикле Кальвина необходимы АТФ и НАДФН. Агранальные хлоропласты клеток обкладки содержат ФС I, поэтому в них происходит только циклическое фотофосфорилирование; это означает, что в этих клетках не восстанавливается НАДФ+. Гранальные хлоропласты мезофильных клеток содержат обе фотосистемы, в них идет циклическое и нециклическое фотофосфорилирование с образованием АТФ и НАДФН.
Когда малат, образованный в мезофильных клетках, поступает в клетки обкладки, то при его декарбоксилировании происходит и окисление, и восстанавливается НАДФ+, необходимый для восстановления ФГК.
Таким образом, С4-цикл поставляет в цикл Кальвина и водород для восстановления СО2.
Эволюционно С3-цикл появился раньше, чем С4, он есть у водорослей. В древесных растениях цикла Хетча – Слека нет. Это тоже подтверждает, что этот цикл возник позднее.
В заключение отметим, что, циклы Кальвина и Хетча – Слека действуют не изолированно, а строго координировано. Взаимосвязь этих двух циклов получила название «кооперативного» фотосинтеза.
Кислотный метаболизм толстянковых (КМТ или САМ цикл). Мы упоминали, что есть растения суккуленты, у которых устьица закрыты для уменьшения транспирации. Это особенность влияет и на фотосинтез. Эти растения поглощают большие количества углекислого газа ночью, когда устьица широко открыты. Поглощенный СО2 используется, как и у С4-растений, для карбоксилирования ФЕП. При этом образуется ЩУК (фермент ФЕП-карбоксилаза содержится в цитоплазме). ЩУК восстанавливается до малата (НАД-зависимая малатдегидрогеназа). Малат накапливается в вакуоли, поэтому ночью вакуолярный сок становится более кислым. В следующее утро малат из вакуоли транспортируется в цитоплазму. Тут происходит его окислительное декарбоксилирование при помощи НАДФ-зависимого малик энзима (малатдегидрогеназа), что приводит к образованию пирувата и СО2 и восстановлению НАДФ+. СО2 и НАДФН и используются в цикле Кальвина, который происходит в хлоропластах, а ПВК фосфорилируется с образованием ФЕП. Последний в результате ряда превращений дает ФГК, которая может также поступать в цикл Кальвина и служит источником для образования сахарозы и крахмала (рис. 2.23).
|
Рис. 2.23. Кислотный метаболизм толстянковых |
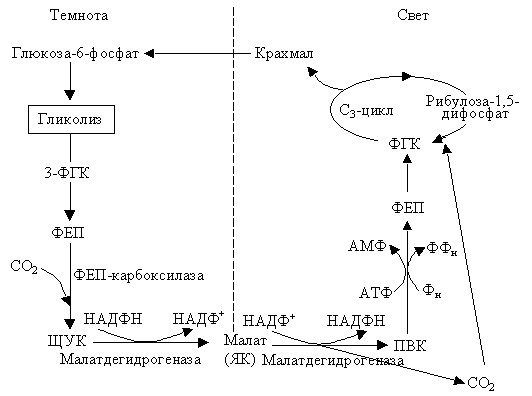
ФЕП, необходимый для акцептирования СО2, образуется ночью в процессе гликолиза.
Следовательно, днем в хлоропластах идет обычный С3-цикл. По мере расходования кислот, накопленных в вакуоли, величина рН в вакуоли снова увеличивается. Для суккулентов – растений сухих мест, разделение во времени ночной фиксации СО2 и переработки СО2 на следующий день выгодно. Это позволяет им осуществлять фотосинтез при закрытых устьицах и одновременно уменьшать транспирацию.
Таким образом, можно отметить, что, для САМ-растений характерны следующие особенности:
– устьица обычно открыты ночью (в темноте) и закрыты на протяжении дня. Такое состояние устьиц противоположно тому, что характерно для других растений;
– фиксация СО2 происходит в темный час суток в содержащих хлоропласты клетках фотосинтезирующих тканей листа или стебля, причем синтезируется значительное количество свободной ЯК;
– ЯК кислота накапливается в больших вакуолях, которые характерны для клеток этих растений;
– на свету ЯК декарбоксилируется и освободившийся СО2 превращается за счет энергии света в сахарозу и запасной глюкан (например, крахмал) в световых реакциях фотосинтеза С3-типа.
– в последующий темновой период часть запасенного глюкана разрушается с образованием молекул-акцепторов для темновой реакции фиксации СО2.
Следовательно, в тканях САМ-растений существует суточный цикл: ночью содержание запасенного глюкана падает и содержание яблочной кислоты повышается, а днем происходят противоположные изменения. Следовательно, реакции разделены во времени, а у растений С4-типа процессы разделены в пространстве (рис. 2.24).
С4-фотосинтез
САМ-фотосинтез
Синтез
СН2О
и запасание
Первичное
карбоксилирование
Синтез
СН2О
Обкладка
сосудистого пучка
Мезофилл
День
Ночь
Пространство
Время
Первичное
карбоксилирование
Рис.
2.24. Схема
разделения во времени и пространстве
фотосинтеза у растений С4-
и САМ-типа

С4 и САМ-типы фиксацин СО2 являются дополнительными, облегчающими функционирование цикла Кальвина в растениях, которые приспособились к жизни в условиях повышенных температур (сахарный тростник, сорго, кукуруза и др.), или недостатка влаги (суккуленты).
В заключении можно дать следующую характеристику процесса фотосинтеза:
– сущность фотосинтеза состоит в фотоокислении воды с выделением кислорода и использованием водорода на восстановление СО2 с образованием органических веществ;
– фотосинтез состоит из двух фаз: световой и темновой;
– энергия поглощенного света используется для движения электронов против градиента окислительно-восстановительного потенциала;
– транспорт электронов сопряжен с синтезом АТФ;
– световая стадия происходит в тилакоидах стромы и гран, а темновая – в строме хлоропластов, т. е. для световой стадии необходимы мембраны.