

Сравнение циклического и нециклического фотофосфорилирования
Процесс |
Нециклическое |
Циклическое |
Путь электронов |
Нециклический |
Циклический |
Первый донор (источник) электронов |
Вода |
ФС I (Р700) |
Последний акцептор (место назначения) электронов |
НАДФ+ |
ФС I (Р700) |
Продукты |
Основные: АТФ, НАДФН Побочные: О2 |
Основные: АТФ |
Участвующие ФС |
I и II |
I |
Электроны, которые поступают от ферредоксина, могут также переноситься и восстанавливать кислород с образованием Н2О2 и в конечном счете Н2О. В этом случае функционируют ФС I и ФС II, и поглощаются с использованием восстановленного ферредоксина в особой реакции равные количества кислорода; такой поток электронов получил название псевдоциклического. Хотя при этом потоке электронов отсутствует суммарное изменение состояния кислорода, этот процесс нельзя назвать циклическим, поскольку не происходит циклического переноса электронов по указанному пути.
|
Синтез АТФ. Энергия, выделяемая при движении электронов от Р680 (Е = –0,8) до Р700 (Е = +0,4 В), используется для синтеза АТФ из АДФ и неорганического фосфата (фотофосфорилирование).
Существуют несколько теорий, разъясняющих механизм фосфорилирования АДФ, сопряженный с работой электрон-транспортной цепи. Наибольшее признание в настоящее время получила хемиосматическая теория английского биохимика П. Митчелла (1961), которая для объяснения процессов фотосинтетического фосфорилирования впервые была использована А. Ягендорфом (1967).
Опыты А. Ягендорфа состояли в следующем (рис. 2.16). Изолированные из хлоропластов граны с рН 8 помещали в буферный раствор с рН 4. После выдерживании в этом растворе в гранах восстанавливалась величина рН, равная 4. Затем граны с рН 4 переносились в буферный раствор с рН 8. Таким образом, на тилакоидных мембранах искусственно создавался градиент протонов, который обычно формируется при фотохимических процессах (внутри тилакоида – рН 4, снаружи – рН 8). В этих условиях после добавления в среду АДФ и Фн в темноте синтезировалось АТФ.
|
Рис. 2.16. Схема эксперимента Ягендорфа |
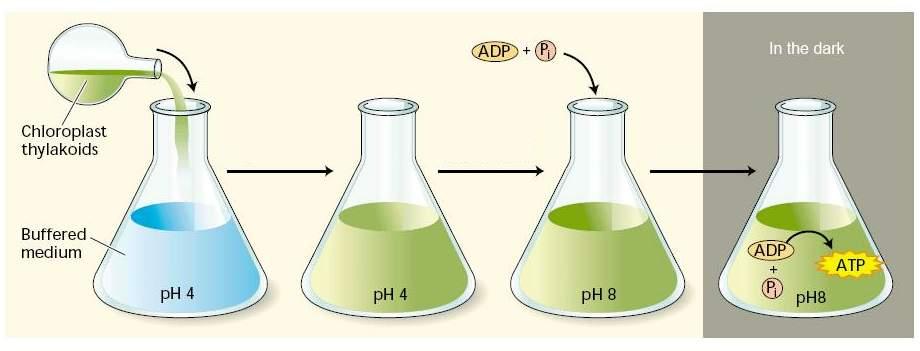
Эти эксперименты позволили применить хемиосмотическую теорию Митчелла для объяснения процессов происходящих при фотофосфорилировании в хлоропластах.
В соответствии с этой теорией пластохинон, присоединивший два электрона, присоединяет еще 2Н+ протона со стороны стромы хлоропласта и переносит их через мембрану во внутритилакоидное пространство. Протоны накапливаются внутри тилакоида и в результате фотоокисления воды.
Б
 лагодаря
неравномерному распределению протонов
по обеим сторонам мембраны создается
разность химических потенциалов ионов
водорода и возникает электрохимический
мембранный потенциал ионов Н+
лагодаря
неравномерному распределению протонов
по обеим сторонам мембраны создается
разность химических потенциалов ионов
водорода и возникает электрохимический
мембранный потенциал ионов Н+
включает две составляющие: концентрационную , возникающую в результате неравномерного размещения ионов Н+ по обеим сторонам мембраны, и электрическую (ΔΨ), которая обусловлена мембранным потенциалом (рис. 2.16).
Энергия ΔрН и ΔΨ используется для обратного транспорта протонов из внутритилакоидного пространства в строму хлоропласта по особым каналам. С обратным транспортом протонов и связано фосфорилирование АДФ. Реакцию катализирует Н+-АТФ-синтетаза, состоящая из двух частей: водорастворимой каталитической части, расположенной в строме хлоропласта, и мембранной части. Последняя представляет собой протонный канал, по которому протоны могут возвращаться в строму хлоропласта. Фермент Н+-АТФ-синтетаза может синтезировать АТФ пока двигаются протоны. Протоны двигаются, когда их концентрация во внутритилакоидном пространстве большая.
|
Рис. 2.17. Иллюстрация гипотезы хемиосмотического сопряжения как механизма фотофосфорилирования |
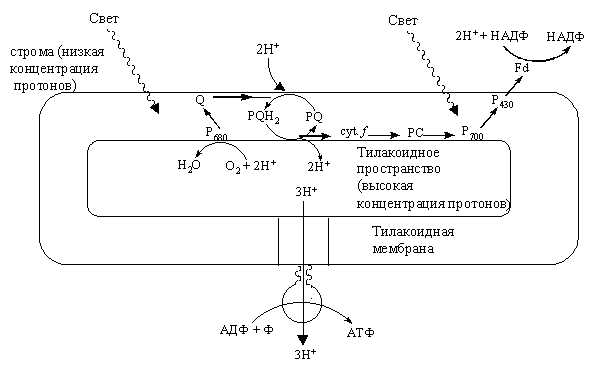
На каждые два электрона, переданных по электрон-транспортной цепи, внутри тилакоида накапливается 4Н+. На каждые 3Н+, возвращающихся назад в строму хлоропласта, синтезируется 1 молекула АТФ.
Таким образом, конечными продуктами световых реакций фотосинтеза является НАДФН и АТФ. Эти соединения используются затем соответственно как восстановительная сила и как источник для превращения СО2 в сахар. Этапы, из которых складываются эти превращения, известны под общим названием «темновых реакций» фотосинтеза.