

2.2.Механизм действия гидрофобных гормонов
Липофильные гормоны ( стероиды ), легко проходят через клеточную мембрану, связываются с рецептором, локализованном либо в цитоплазме, либо в ядре клетки.
Стероидные гормоны и тиреоидные гормоны имеют очень ограниченную растворимость в воде и для них физически невозможно существование в виде свободного раствора при их известных концентрациях в плазме. При этих концентрациях они циркулируют только потому, что существует группа плазменных белков, которые связывают эти гормоны, удаляя их из свободного раствора, но сохраняя их в плазме ( рис.1-7сер ). Некоторые из этих белков являются высоко специализированными, они связывают только определенные гормоны и, по-видимому, не имеют никаких других функций. Другие, наоборот, связывают все гидрофобные гормоны. Степень связывания гормона с белком и распределение гормона между различными гормонсвязывающими белками варьирует у разных гормонов.
Однако, во всех случаях суммарный гормональный пул представлен преимущественно связанной формой; в некоторых случаях только 1% или меньше суммарного гормона остается в несвязанном с белком состоянии. Эти цифры заслуживают внимания, так как существует общее мнение, что только свободный гормон является биологически активной формой гормона. Следует отметить, что гормон, связанный с белком-носителем, находится в динамическом равновесии с пулом свободного гормона, который в свою очередь находится в равновесии с гормоном, связанным с рецепторами в органе-мишени. Тем не менее, поскольку гормон связан с белком плазмы, он не может взаимодействовать со своим рецептором и таким образом является неактивным. Однако, очевидно, что помимо облегчения растворения гидрофобных гормонов в плазме, связывающие белки формируют большой гормональный резерв, который при необходимости может быть быстро использован для пополнения пула свободного гормона. Размеры свободного пула в большей степени, чем общий пул гормона обеспечивают поддержание нормальной эндокринной функции.
Величина свободного пула и, таким образом, доступность гормона рецептору может регулироваться изменениями секреции, метаболизма или белкового связывания. Легко понять, что концентрация гормона в плазме будет находиться в прямой зависимости от скорости его секреции. Другие факторы, влияющие на концентрацию свободного гормона, могут быть трудноуловимы, но они также достаточно важны как с познавательной, так и с клинической точки зрения. Например, величина свободного пула будет иметь тенденцию к увеличению при любых состояниях, снижающих скорость обмена гормона (деградацию). Существуют такие физиологические (беременность) и патофизиологические состояния (болезнь печени), при которых происходят изменения баланса свободного и связанного пулов определенных гормонов. С диагностической точки зрения важно понимать, что используемые методы определения этих гормонов позволяют измерять суммарную их концентрацию, т.е. сумму свободного и связанного с белками гормона. В этих случаях, поэтому, необходимо также проводить некое определение размеров связывания гормона с белком, с тем чтобы определить концентрацию свободного гормона.
Гормоны, поступая в кровь, транспортируются ею к органам и тканям. При этом лишь небольшая часть гормона циркулирует в свободной форме в виде водного раствора. Преобладающая же часть его находится в крови в форме обратимых комплексов с белками плазмы и форменными элементами . В этих комплексах составляющие компоненты сцеплены нековалентными связями. Особое значение имеют транспортные белки плазмы , специфически связывающие те или иные гормоны. К специфически связывающим транспортным белкам от- носятся такие белки, кактранскортин , избирательно связывающий кортикостероиды и прогестины , секс-стероидсвязывающий глобулин , образующий комплексы с андрогенами и эстрогенами ,тироксинсвязывающий глобулин , образующий комплексы с тиреоидными гормонами , и т.д. Вступая в комплекс с этими белками, гормоны аккумулируются в кровяном русле и, тем самым, временно выключаются из сферы биологического действия и метаболических превращений, т.е. обратимо инактивируются. Активной становится свободная, несвязанная с белками форма. По-видимому, образование комплексов гормонов со специфическими белками плазмы есть механизм буферирования сдвигов концентрации гормонов и их резервирования в кровяном русле.
Стероидные гормоны - один из главных классов гормональных соединений всех видов позвоночных и многих видов беспозвоночных животных. Они являются регуляторами фундаментальных процессов жизнедеятельности многоклеточного организма - координированного роста , дифференцировки , размножения , адаптации , поведения [Хартман Э., 1972].
Действие стероидных гормонов на клетки-мишени осуществляется, главным образом, на уровне регуляции транскрипции генов . Оно опосредуется образованием комплекса гормона со специфическим регуляторным белком - рецептором , узнающим определенные участки ДНК в генах, регулируемых данным гормоном. Таким образом, рецепторы всех стероидных гормонов - лиганд-зависимые факторы транскрипции. Для них характерно значительное сходство аминокислотных последовательностей, идентичная доменная структура и сходный механизм действия. Вместе с близкими им рецепторами тиреоидных гормонов они объединяются в семейство рецепторов стероидных/тиреоидных гормонов , которое входит в более обширное надсемейство регуляторных белков - ядерных рецепторов [ Mangelsford D.J. et al., 1995 ].
Стероидные гормоны синтезируются из холестерина, в основном, в коре надпочечников, тестикулах, яичниках и плаценте; однако, каждая ткань, продуцирующая стероиды, имеет свой собственный характерный профиль продуктов секреции.
Подобно надпочечникам, половые железы продуцируют довольно много стероидов, но лишь некоторые из них обладают гормональной активностью. Образование этих гормонов строго регулируется с помощью петли обратной связи , включающей в себя гипофиз и гипоталамус . Действие половых гормонов опосредовано ядерными механизмами, подобными тем, которые используются кортикостероидами.
Стероидные соединения плохо растворяются в воде и хорошо в органических растворителях и растительных маслах. Все стероидные гормоны благодаря липофильности относительно легко проникают через плазматические липопротеидные мембраны и поэтому могут свободно секретироваться клетками стероидогенных эндокринных желез и входить внутрь реагирующих клеток.
стероиды характеризуются трехмерной пространственной конфигурацией, особенности которой оказывают существенное влияние на их биологическую активность.
Гормоны этого класса представляют полициклические соединения липидной природы, в основе структуры которых, как правило, лежит циклопентанпергидрофенантреновое (стерановое) ядро, состоящее из конденсированных между собой трех насыщенных шестичленных (А,В,С) и одного насыщенного пятичленного кольца (D).
Структурно стероиды отличаются друг от друга количеством и расположением функциональных групп, степенью насыщенности углеводных связей, длиной боковой цепи, прикрепленной к стероидному ядру, и другими химическими характеристиками. Структуры некоторых физиологически важных стероидных гормонов представлены на рис.2-3 .
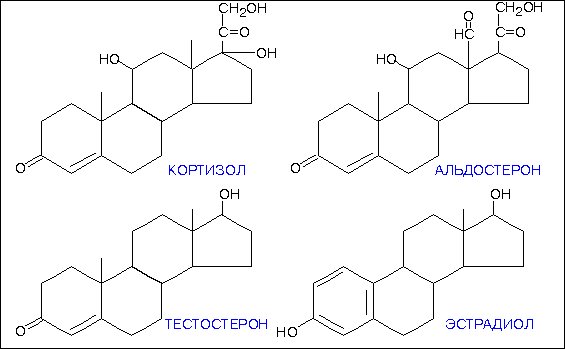
Структура основных стероидных гормонов.
Хотя часто трудно установить определенную взаимосвязь между структурой и активностью, определенные структурные черты, по всей вероятности, являются общими для стероидов со сходной биологической активностью. Например, стероиды с андрогенной или маскулинизирующей активностью, типичным представителем которых является тестостерон , содержат 19 углеродных атомов и не имеют боковой цепи, прикрепленной к стероидному ядру (рис.2-3 ). Соединения с эстрогенной или феминизирующей активностью, такие как эстрадиол , содержат 18 углеродных атомов и имеют ароматическое кольцо А. Другие физиологически важные стероидные гормоны содержат 21 углеродный атом и включают в себя кортикостероиды и прогестины.
Геометрическая форма стероидной молекулы достаточно стабильна и определяется конформацией колец (циклогексановые кольца имеют конформацию кресла, а циклопентановое - полукресла), характером их соединения (В/С- и С/D-сочленения имеют транс-конфигурацию), а также расположением атомов водорода, радикалов и функциональных групп, присоединенных к стериновому ядру (альфа-или бета-ориентация в пространстве). Следует отметить, что стероиды - оптически активные соединения. Все природные гормоны являются производными D-ряда.
класс стероидных гормонов включает семейства производных четырех углеродных стероидов, содержащих различное количество углеродных атомов (С*4n*5): * прегнана (С*421*5),андростана (С*419*5), эстрана (С*418*5) и холестана (С*427*5).
Прегнановые стероидные гормоны *можно рассматривать как производные С*421*стероида прегнана. Это семейство гормонов состоит из двух основных групп: кортикостероидов - гормонов коры надпочечников и прогестинов *- гормонов яичников и плаценты