

Строение
Границей между мостом и продолговатым мозгом с дорсальной стороны являются мозговые полоски, с вентральной — поперечная борозда; между мостом и средним мозгом — место отхождения IV (блокового) черепномозгового нерва.С вентральной (передней) стороны мост представляет собой валик с продольным центральным углублением — базиллярной бороздой, sulcus basillaris, в которой лежат артерии, снабжающие головной мозг. Латеральнее ее в середине валика мост покидает самый толстый, V (тройничный) черепномозговой нерв.С дорсальной стороны мост представлен верхней половиной ромбовидной ямки. Над медуллярными полосками представлены лицевые холмики, colliculus faciales. В них залегают ядра VI (отводящего) нерва, вокруг которых делает петлю VII нерв — так называемое колено лицевого нерва. Выше лицевых холмиков — срединное возвышение, eminentia medialis. Латеральнее его — голубое пятно, часть ретикулярной формации.От моста отходят (из боковых углов ромбовидной ямки) средние ножки мозжечка, они же ручки моста, brachii ponti — толстые проводящие пути. Несколько выше их — верхние ножки мозжечка, они же соединительные ручки, brachium conjuctivum.В варолиевом мосту в его покрышке (верхней части) залегают ядра следующих нервов: частично — VIII, два ядра VII нерва, VI и V (одно из ядер уходит ниже, еще одно — выше). Также в нем содержится часть ретикулярной формации, продолжение медиальной петли, относящиеся к слуховым путям трапециевидное тело, ядра верхней оливы и латеральная петля.В базисе моста проходят нисходящие пути: кортикоспинальный пирамидный тракт, кортикобульбарный, кортикомостовый тракты.]Функции
Здесь находится центр дыхания
Передача информации из спинного мозга в отделы головного
17.Строение мозжечка
Мозжечок (cerebellum) расположен под дубликатурой твердой мозговой оболочки, называемой наметом мозжечка (tentorium cerebelli). Намет делит полость черепа на неравные пространства — субтенториальное и супратенториальное. Дном субтенториального пространства является задняя черепная ямка. В намете, помимо мозжечка, располагается ствол мозга. В среднем объем мозжечка — около 160 см3. Масса, как правило, составляет 140—170 г.

Местоположение мозжечка - над продолговатым мозгом и мостом. Вместе с мозговыми парусами (верхним и нижним) он образует крышу IV желудочка мозга. Так называемая ромбовидная ямка является дном этого желудочка. Над мозжечком расположены затылочные доли большого мозга, которые отделены от него наметом мозжечка. Мозжечок делится на два полушария (hemispherum cerebelli), между которыми над IV желудочком мозга в сагиттальной плоскости находится наиболее древняя, с филогенетической точки зрения, часть мозжечка. Это его червь (vermis cerebelli), который вместе с полушариями мозжечка глубокими поперечными бороздами фрагментируются на дольки. Составляют мозжечок серое и белое вещества. Из серого вещества формируется кора мозжечка и парные ядра (nuclei cerebelli), находящиеся в его глубине. Самыми крупными из них являются зубчатые ядра (nucleus dentatus), они расположены в полушариях. Центральная часть червя имеет ядра шатра (nucleifastigii). Между этими и зубчатыми ядрами расположены пробковидные и шаровидные ядра (nuclei, emboliformis et globosus). На сагиттальном разрезе мозжечка, в результате того, что корой покрывается вся поверхность мозжечка, проникающая в глубину его борозд, его ткань имеет рисунок листа с прожилками, образованными белым веществом. Эти прожилки составляют так именуемое древо жизни мозжечка (arbor vitae cerebelli). В его основании расположена клиновидная выемка, которая является верхней частью полости IV желудочка. Краями этой выемки формируется шатер. Червь мозжечка служит крышей этого шатра, а тонкие мозговые пластинки составляют его переднюю и заднюю стенки, называемые передним и задним мозговыми парусами (vella medullare anterior et posterior). У мозжечка имеется три пары ножек: нижняя, средняя и верхняя. С помощью них осуществляются связи мозжечка с продолговатым мозгом, мостом и со средним мозгом. Некоторые из сведений об архитектонике мозжечка представляют немалый интерес, они позволяют судить о функции его компонентов. Кора мозжечка имеет два клеточных слоя: наружный — молекулярный и внутренний — зернистый, который состоит из мелких клеток-зерен. Между слоями располагается ряд крупных грушевидных клеток, которые своим названием обязаны описавшему их чешскому ученому И. Пуркинье (Purkinje I., I787-1869).
18.Строение среднего мозга
Средний мозг располагается между задним и промежуточным мозгом. Он состоит из двух основных отделов: крыши среднего мозга и ножек большого мозга (см. рис.25, 26, 29). Границей между ними считается плоскость, проходящая параллельно пластинке четверохолмия через водопровод мозга. Водопровод мозга является полостью среднего мозга, он представляет собой узкий канал длиной около 1,5 см, соединяющий полости III и IV желудочков.
Крыша среднего мозга, или пластинка четверохолмия является дорсальной частью среднего мозга (см. рис. 29). На целом мозге она не видна, так как сверху над ней нависает задняя часть мозолистого тела и затылочные доли полушарий большого мозга, а снизу – верхняя часть мозжечка (см. рис. 25). На наружной поверхности крыши среднего мозга располагаются четыре возвышения, бугорка – холмики. Различают верхнее и нижнее двухолмие, каждое состоит из правого и левого холмика. Верхние холмики крупнее нижних.
В верхних холмиках располагаются подкорковые центры (переключающие ядра) зрительного анализатора, а в нижних – слухового анализатора. По бокам от каждого холмика к промежуточному мозгу отходят ручки холмиков (см. рис. 29). Ручка верхнего холмика направляется к латеральному коленчатому телу (СНОСКА: Латеральное коленчатое тело – структура заталамической области (см. Раздел 6.4.3.), является подкорковым центром зрительного анализатора), а ручка нижнего холмика – к медиальному коленчатому телу (СНОСКА: Медиальное коленчатое тело также относится к промежуточному мозгу (см. Раздел 6.4.3.), является подкорковым центром слухового анализатора).
Ножки мозга располагаются вентральнее крыши среднего мозга. Ножки мозга выглядят в виде толстых округлых тяжей белого цвета, выходящих из моста и направляющихся вперёд, вверх и немного в стороны друг от друга к полушариям большого мозга (см. рис. 26, 27). Между ножками находится межножковая ямка, на дне которой видно заднее продырявленное вещество. На медиальной поверхности каждой ножки выходит глазодвигательный нерв (III пара).
На фронтальных разрезах ножек мозга различают две части (рис. 33):вентральную (основание ножки мозга) и дорсальную (покрышку ножки мозга). Границей между ними является чёрное вещество, состоящее из клеток, содержащих темный пигмент меланин.
Основания ножек мозга образованы нервными волокнами двигательных пирамидных путей, идущими от коры большого мозга к двигательным ядрам моста, продолговатого и спинного мозга.
Покрышки ножек мозга содержат, главным образом, восходящие (чувствительные) проводящие пути, направляющиеся к таламусу, а также крупные и мелкие скопления нейронов – ядра покрышки среднего мозга. Между покрышками правой и левой ножек мозга, медиально, располагаются перекрёсты покрышки. Они образованы пучками волокон красноядерно-спинномозгового, красноядерно-ретикулярного и покрышечно-спинномозгового путей, переходящих на уровне среднего мозга на противоположную сторону.
Большинство ядер покрышки (кроме срединного) являются парными – по одному в каждой покрышке ножек среднего мозга. Самыми крупными из ядер покрышки являются: 1) красные ядра, расположенные в центре покрышек, 2) ядра глазодвигательных нервов (III пара), расположенные дорсальнее красных ядер. Вокруг водопровода мозга располагаются 3) ядра блоковых нервов (IV пара), 4) парное добавочное вегетативное (Якубовича), 5) ядра среднемозгового пути тройничного нерва (V пара), 6) промежуточные ядра дорсального продольного пучка и 7) ретикулярная формация.
19 Строение промежуточного мозга – таламическая область
22 Строение большого мозга
Конечный мозг (telencephalon), или большой мозг (cerebrum), располагается в супратенториальном пространстве полости черепа и состоит из двух больших полушарий (gemispherium cerebralis), разделенных между собой глубокой продольной щелью (fissura longiiudinalis cerebri), в которую погружен серп большого мозга ifalx cerebri), представляющий собою дубликатуру твердой мозговой оболочки. Большие полушария головного мозга составляют 78% его массы. Каждое из больших полушарий имеет доли: лобную, теменную, височную, затылочную и лимбическую. Они прикрывают структуры промежуточного мозга и находящиеся ниже мозжечкового намета (субтенториально) мозговой ствол и мозжечок.
Каждое из полушарий большого мозга имеет три поверхности: верхнебоковую, или конвекситальную (рис. 14.1а), — выпуклую, обращенную к костям свода черепа; внутреннюю (рис. 14.16), прилежащую к большому серповидному отростку, и нижнюю, или базальную (рис. 14.1в), повторяющую рельеф основания черепа (передней и средней его ямок) и намета мозжечка. В каждом полушарии различают три края: верхний, нижневнутренний и нижненаружный, и три полюса: передний (лобный), задний (затылочный) и боковой (височный).
Полость каждого большого полушария составляет боковой желудочек мозга, при этом левый боковой желудочек признается первым, правый — вторым. Боковой желудочек имеет центральную часть, находящуюся в глубине теменной доли (lobus parietalis) и три отходящих от нее рога: передний рог проникает в лобную долю (lobus frontalis), нижний — в височную (lobus temporalis), задний — в затылочную (lobus occipitalis). Каждый из боковых желудочков сообщатся с третьим желудочком мозга через межжелудочковое отверстие Монро.
Центральные отделы медиальной поверхности обоих полушарий соединены между собой мозговыми спайками, наиболее массивной из которых является мозолистое тело, и структурами промежуточного мозга.
23 Особенности строения коры большого мозга
.Корковые структуры
Кора больших полушарий головного мозга, слой серого вещества толщиной 1—5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга, развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности, хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468—1670 см2. Строение коры. Характерной особеннос тью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними (рис. 1). Пространство между телами и отростками нервных клеток коры заполнено нейроглией и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80—90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры — афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон (рис. 2). Звездчатые клетки отличаются слабым развитием дендритов и мощным развитием аксонов, которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением (см. Клетка). Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или её цитоархитектонику
Корковые структуры
Кора покрывает всю поверхность больших полушарий. Ее структурными элементами являются нервные клетки с отходящими от них отростками - аксонами и дендритами - и клетки нейроглии.
В коре полушарий большого мозга человека насчитывают около 12-18 млрд. нервных клеток. Из них 8 млрд. составляют крупные и средних размеров клетки третьего, пятого и шестого слоев, около 5 млрд. приходится на мелкие клетки различных слоев. [показать]
Основная масса клеток коры состоит из элементов трех родов:
пирамидных клеток
веретенообразных клеток
звездчатых клеток
Полагают, что пирамидные и веретенообразные клетки с длинными аксонами представляют преимущественно эфферентные системы коры, а звездчатые - преимущественно афферентные. Считают, что клеток нейроглии в головном мозге в 10 раз больше, чем ганглиозных (нервных) клеток, т. е. около 100-130 млрд. Толщина коры варьирует от 1,5 до 4 мм. Общая поверхность обоих полушарий коры у взрослого человека составляет от 1450 до 1700 см2.
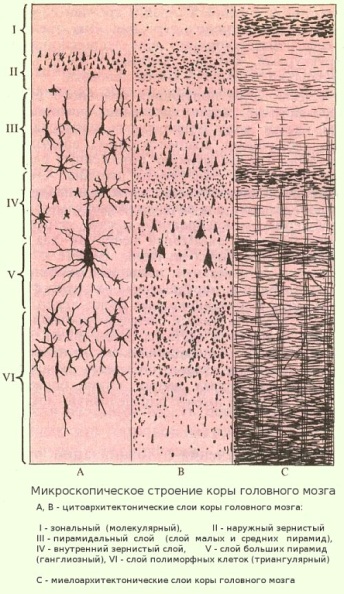
Особенностью структуры коры больших полушарий является расположение нервных клеток в шесть слоев, лежащих друг над другом.
первый слой - lamina zonalis, зональный (краевой) слой или молекулярный - беден нервными клетками и образован в основном сплетением нервных волокон
второй - lamina granularis externa, наружный зернистый слой - называется так из-за наличия в нем густо расположенных мелких клеток, диаметром 4-8 мк,имеющих на микроскопических препаратах форму круглых, треугольных и многоугольных зерен
третий - lamina pyramidalis, пирамидальный слой - имеет большую толщину, чем первые два слоя. В нем содержатся пирамидные клетки разной величины
четвертый - lamina dranularis interna, внутренний зернистый слой - подобно второму слою, он состоит из мелких клеток. Этот слой в некоторых участках коры больших полушарий взрослого организма может отсутствовать; так, например, его нет в моторной области коры
пятый - lamina gigantopyramidalis, слой больших пирамид (гигантские клетки Беца) - от верхней части этих клеток отходит толстый отросток - дендрит, многократно ветвящийся в поверхностных слоях коры. Другой длинный отросток - аксон - больших пирамидных меток уходит в белое вещество и направляется к подкорковым ядрам или к спинному мозгу.
шестой - lamina multiformis, полиморфный слой (мультиформный) - состоит из клеток треугольной формы и веретенообразных
По функциональному признаку нейроны коры больших полушарий могут быть подразделены на три основные группы.
Сенсорные нейроны коры больших полушарий, так называемые звездчатые нейроны, которые в особенно большом количестве находятся в III и IV слоях сенсорных областей коры. На них оканчиваются аксоны третьих нейронов специфических афферентных путей. Эти клетки обеспечивают восприятие афферентных импульсов, приходящих в кору больших полушарий из ядер зрительных бугров.
Моторные (эффекторны) нейроны - клетки, посылающие импульсы в лежащие ниже отделы мозга - к подкорковым ядрам, стволу мозга и спинному мозгу. Это большие пирамидные нейроны, которые впервые описал В. А. Бец в 1874 г. Они сконцентрированы в основном в V слое моторной зоны коры. В осуществлении эффекторной функции коры принимают участие и некоторые веретенообразные клетки.
Контактные, или промежуточные, нейроны - клетки, осуществляющие связь между различными нейронами одной и той же или различных зон коры. К их числу относятся мелкие и средние пирамидные и веретенообразные клетки.
24 Локализация функций в коре головного мозга
Вместо представления о коре больших полушарий как, в известной мере, изолированной надстройке над другими этажами нервной системы с узко локализованными, связанными по поверхности (ассоциационными) и с периферией (проекционными) областями, И.П. Павлов создал учение о функциональном единстве нейронов, относящихся к различным отделам нервной системы — от рецепторов на периферии до коры головного мозга — учение об анализаторах. То, что мы называем центром, является высшим, корковым, отделом анализатора. Каждый анализатор связан с определенными областями коры головного мозга. И.П. Павлов вносит существенные коррективы в прежние представления об ограниченности территорий корковых центров, в учение об узкой локализации функций. Вот что говорит он о проекции рецепторов в кору головного мозга. «Каждый периферический рецепторный аппарат имеет в коре центральную, специальную, обособленную территорию, как его конечную станцию, которая представляет его точную проекцию. Здесь благодаря особенной конструкции, может быть более плотному размещению клеток, более многочисленным соединениям клеток и отсутствию клеток других функций, происходят, образуются сложнейшие раздражения (высший синтез) и совершается их точная дифференцировка (высший анализ). Но данные рецепторные элементы распространяются и дальше на очень большое расстояние, может быть по всей коре». С этим выводом, основанном на обширных экспериментально-физиологических исследованиях, вполне согласуются новейшие морфологические данные о невозможности точного разграничения корковых цитоархитектонических полей. Следовательно, функции анализаторов (или, иными словами, работу первой сигнальной системы) нельзя связывать только с корковыми проекционными зонами (ядрами анализаторов). Тем более нельзя узко локализовать сложнейшие, чисто человеческие функции — функции второй сигнальной системы. И.П. Павлов следующим образом определяет функции сигнальных систем человека. «Всю совокупность высшей нервной деятельности я представляю себе так. У высших животных, до человека включительно, первая инстанция для сложных соотношений организма с окружающей средой есть ближайшая к полушариям подкорка с ее сложнейшими безусловными рефлексами (наша терминология), инстинктами, влечениями, аффектами, эмоциями (разнообразная, обычная терминология). Вызываются эти рефлексы относительно немногими безусловными внешними агентами. Отсюда — ограниченная ориентировка в окружающей среде и вместе с тем слабое приспособление. Вторая инстанция — большие полушария. Тут возникает при помощи условной связи (ассоциации) новый принцип деятельности: сигнализация немногих, безусловных внешних агентов бесчисленной массой других агентов, постоянно вместе с тем анализируемых и синтезируемых, дающих возможность очень большой ориентировки в той же среде и тем же гораздо большего приспособления. Это составляет единственную сигнализационную систему в животном организме и первую в человеке.
25. Общая характеристика базальных ядер
Базальные ядра обеспечивают двигательные функции,.отличные от таковых, контролируемых пирамидным (кортико-спинальным) трактом. Термин экстрапирамидный подчеркивает это различие и относится к ряду заболеваний, при которых поражаются базальные ядра. К семейным заболеваниям относят болезнь Паркинсона, хорею Гентингтона и болезнь Вильсона. В этом параграфе рассматривается вопрос о базальных ядрах и описываются объективные и субъективные признаки нарушений их деятельности.
Анатомические связи и нейротрансмиттеры базальных ядер.Базальные ядра представляют собой парные подкорковые скопления серого вещества, образующие обособленные группы ядер. Основными являются хвостатое ядро и скорлупа (вместе формирующие полосатое тело), медиальная и латеральная пластинки бледного шара, субталамическос ядро и черное вещество (рис. 15.2). Полосатое тело получает афферентные сигналы из многих источников, включая кору больших полушарий, ядра зрительного бугра, ядра шва ствола мозга и черное вещество. Корковые нейроны, связанные с полосатым телом, выделяют глутаминовую кислоту, обладающую возбуждающим эффектом. Нейроны ядер шва, связанные с полосатым телом, синтезируют и выделяют серотонин. (5-ГТ). Нейроны компактной части черного вещества синтезируют и выделяют дофамин, который воздействует на нейроны полосатого тела в качестве тормозного медиатора. Трансмиттеры, выделяемые проводниками таламуса, не определены. Полосатое тело содержит 2 вида клеток: местные обходные нейроны, аксоны которых не выходят за пределы ядер и остальные нейроны, аксоны которых идут к бледному шару и черному веществу. Местные обходные нейроны синтезируют и выделяют ацетилхолин, гамма-аминомасляную кислоту (ГАМК) и нейропептиды, такие как соматостатин и вазоактивный интестинальный полипептид. Нейроны полосатого тела, обладающие подавляющим воздействием на ретикулярную часть черного вещества, выделяют ГАМК, тогда как те, которые возбуждают черное вещество, выделяют субстанцию Р (рис. 15.3). Стриарные проекции к бледному шару выделяют ГАМК, энкефалины и субстанцию Р.
26.Общая характеристики ратикулярной формации
Ретикулярная формация - это продолговатая структура в стволе мозга . Она представляет собой важный пункт на пути восходящей неспецифической соматосенсорной системы .Соматовисцеральные афференты идут в составе спиноретикулярного тракта ( переднебоковой канатик ), а также, возможно, в составе проприоспинальных (полисинаптических) путей и соответствующих путей от ядра спинального тройничного тракта . К ретикулярной формации приходят также пути от всех других афферентных черепномозговых нервов , т.е. практически от всех органов чувств. Дополнительная афферентация поступает от многих других отделовголовного мозга - от моторных областей коры и сенсорных областей коры , от таламуса игипоталамуса . Имеется также множество эфферентных связей - нисходящие к спинному мозгуи восходящие через неспецифические таламические ядра к коре головного мозга ,гипоталамусу и лимбической системе . Большинство нейронов образует синапсы с двумя - тремя афферентами разного происхождения, такая полисенсорная конвергенция характерна для нейронов ретикулярной формации. Другими их свойствами являются большие рецептивные поляповерхности тела, часто билатеральные, длительный латентный период ответа на периферическую стимуляцию (вследствие мультисинаптического проведения), слабая воспроизводимость реакции (стохастические колебания числа потенциалов действия при повторной стимуляции). Все эти свойства противоположны свойствам лемнисковых нейронов вспецифических ядрах соматосенсорной системы ( рис.9-7 и рис. 5-13 ).
Функции ретикулярной формации изучены не полностью. Считается, что она участвует в следующих процессах:
1. в регуляции уровня сознания путем воздействия на активность корковых нейронов , например, участие в цикле сон / бодрствование ,
2. в придании аффективно-эмоциональной окраски сенсорным стимулам, в том числе болевым сигналам , идущим по переднебоковому канатику , путем проведения афферентной информации к лимбической системе ,
3. в вегетативных регулирующих функциях, в том числе во многих жизненно важных рефлексах ( циркуляторных рефлексах и дыхательных рефлексах , рефлекторных актах глотания , кашля ,чихания ), при которых должны взаимно координироваться разные афферентные и эфферентные системы,
4. в целенаправленных движениях в качестве важного компонента двигательных центров ствола мозга .
27.Общая характеристика лимбической системы
Лимбическая система (от латинского limbus - кайма) - обширная нейронная структура - является морфофункциональным комплексом структур, которые расположены в различных отделах конечного мозга и промежуточного мозга (рис. 68 ). Лимбическую систему формируют лимбические и паралимбические структуры (ряд образований на медиальной и нижней поверхностях полушарий продолговатого мозга, передние и медиальные ядра таламуса , медиальные и базальные отделы стриатума, а также гипоталамус) (табл. 25.1). Она координирует эмоциональные, мотивационные, вегетативные и эндокринные процессы. В нее включены древние подкорковые и плащевые структуры. От конечного мозга в нее входят поясная извилина , зубчатая извилина , гиппокамп (морской конек), септум (перегородка) и миндалевидные тела . В промежуточном мозге расположены 4 основные структуры лимбической системы: хабенулярные ядра (ядра поводков) , таламус , гипоталамус и сосцевидные тела . Волокна, соединяющие структуры лимбической системы, образуют свод конечного мозга , который проходит в виде арки от архикортекса до сосцевидных тел . Лимбическая система объединена многочисленными связями с неокортексом и автономной нервной системой, поэтому она интегрирует две важнейшие функции мозга животного и человека - эмоции и память . Удаление части лимбической системы приводит к эмоциональной пассивности животного, а стимуляция - к эмоциональной гиперактивности . Активизация миндалевидного комплекса запускает механизмы агрессии , которые могут корректироватьсягиппокампом . Лимбическая система запускает пищевое поведение и вызывает чувство опасности . Все эти формы поведения контролируются как самой лимбической системой, так и через гормоны, вырабатываемые гипоталамусом . Влияние лимбической системы на функции организма осуществляется через контроль за деятельностью автономной нервной системы. Роль лимбической системы столь высока, что ее называют висцеральным мозгом. Она обусловливает эмоционально- гормональную активность животного, которая, как правило, плохо поддается рассудочному контролю даже у человека.Важнейшей функцией лимбической системы является взаимодействие с механизмами памяти. Краткосрочную память обычно связывают с гиппокампом , адолгосрочную - с неокортексом . Однако извлечение индивидуального опыта животного и человека из неокортекса осуществляется через лимбическую систему. При этом используется эмоционально-гормональная стимуляция мозга, которая вызывает информацию из неокортекса.Лимбическая система обеспечивает и еще одну важную функцию, нарушение которой часто встречается в клинической практике, - декларативную, или вербальную, память о событиях, приобретенных навыках и накопленных знаниях.Лимбическая система обладает уникальным набором эффекторных структур. В них входят управление моторикой внутренних органов, двигательная активность для выражения эмоций и гормональная стимуляция организма . Чем ниже уровень развития неокортекса , тем большеяповедение животного зависит от лимбической системы.После предварительной переработки в центральной нервной системе второй уровень центральной переработки информации происходит в четырех функциональных системах, к которым приходят сигналы от органов чувств. Это ассоциативная система, лимбическая система , двигательная система ивегетативная система . Сложное, почти неисследованное взаимодействие этих областей мозга можно считать основой нашего поведения. В основном к лимбической системе относят структуры обонятельного мозга - наиболее древней части полушарий. В описаниях морфологов лимбическую систему представляют в виде "анатомического эмоционального кольца", в состав которого входят различные образования мозга ( рис. 223 ). Это корковые структуры: гиппокамп , парагиппокампова извилина , поясная извилина , структуры обонятельного мозга (обонятельные луковицы , обонятельные бугорки ), области коры над миндалиной, а также частично кора лобной , кора островковой и кора височной долей; подкорковые структуры ( миндалина , ядра перегородки, ядра таламуса передние ), гипоталамус, сосцевидные тела . Как уже было сказано выше, все лимбические структуры связаны между собой и с другими отделами мозга. Особенно богаты связи с гипоталамусом. Кора лобных долей регулирует деятельность лимбической системы. Через лимбическую систему проходят сигналы, направляющиеся от всех органов чувств в кору полушарий , а также в обратном направлении. Она обусловливает эмоциональный настрой человека и мотивации, т. е. побуждение к действию, поведение, процессы научения и памяти, а также обеспечивает общее улучшение приспособления организма к постоянно изменяющимся условиям внешней среды.Несмотря на то что поражение лимбических структур вызывает амнезию , лимбическую систему нельзя считать хранилищем. Следы памяти распределены по всей ассоциативной коре , и роль лимбической системы состоит в объединении этих отдельных фрагментов в доступные для припоминания события и знания. Поражение лимбической системы не стирает следы памяти, а нарушает их сознательное воспроизведение, при этом отдельные фрагменты информации остаются сохранными и обеспечивают так называемую процедурную память . Так, больные с корсаковским синдромом способны усвоить новый двигательный или перцептивный навык, но при этом не помнят, как и чему научились.
28.Общая характеристика черепных нервов
ЧЕРЕПНЫЕ НЕРВЫ: ХАРАКТЕРИСТИКИ [ cranial nerves: characteristics ]
Черепные нервы представляют собой двенадцать пар нервов, происходящих из нервной ткани головного мозга. Их обозначают римскими цифрами по порядку их расположения. Черепные нервы входят (афферентные, чувствительные волокна) или выходят (эфферентные, соматические и вегетативные волокна) в/из головного мозга в определенных местах, через особые отверстия в черепе (см. рисунки в списке литературы 6). Назначение любых нервов, в том числе и черепных нервов, проводить как афферентную, так и эфферентную информацию. Афферентная информация передается по афферентным нервным волокнам. Эти волокна являются отростками афферентных (чувствительных) нейронов, расположенных в афферентных узлах (ганглиях), как правило, вне центральной нервной системы. Афферентная информация проходит по афферентным волокнам нервов от рецепторов различных структур организма (тканей,органов и систем) к регуляторам центральной нервной системы - нервным центрам. Ядра центральной нервной системы являются компонентами таких центров. Афферентная информация используется регуляторами для формирования управляющих сигналов - эфферентной информации, поступающей к объектам управления - различным структурам организма. Эфферентная информация передается по эфферентным волокнам от эфферентных нейронов нервных центров. Эфферентные нейроны могут быть расположены как в центральной нервной системе (соматические нервные центры, ядра; вегетативные нервные центры, ядра), так и вне центральной нервной системы (вегетативные нервные центры, ядра). Все нервы являются смешанными, то есть могут содержать как афферентные, так и эфферентные соматические или вегетативные волокна. Если в составе нерва преобладает какой-либо один тип волокон, этот нерв имеет соответствующее название. Например, чувствительный нерв, соматический двигательный нерв, эфферентный висцеральный парасимпатический нерв и т.д. В таблице ниже показаны главные характеристики черепных нервов: нервные волокна, составляющие черепные нервы,ядра черепных нервов, области, иннервируемые черепными нервами, назначение и функции черепных нервов. См. также описание некоторых ядер черепных нервов в статье четвертый желудочек.
ЧН I – Обонятельный |
ЧН II – Зрительный |
ЧН III – Глазодвигательный |
ЧН IV – Блоковый |
ЧН V – Тройничный |
ЧН VI – Отводящий |
ЧН VII – Лицевой |
ЧН VIII – Преддверно-улитковый |
ЧН IX – Языкоглоточный |
ЧН X – Блуждающий |
ЧН XI – Добавочный |
ЧН XII – Подъязычный |
29.Обонятельные нервы типография, Области инервации
Обонятельные нервы представляют собой нервы специальной чувствительности — обонятельной. Они начинаются от обонятельных нейросенсорных клеток, образующихпервый нейрон обонятельного пути и залегающих в обонятельной области слизистой оболочки полости носа. В виде 15-20 тонких нервных стволов (обонятельные нити), состоящих из безмиелиновых нервных волокон, они, не образуя общего ствола обонятельного нерва, проникают через горизонтальную пластинку решётчатой кости(лат. lamina cribosa ossis ethmoidalis) в полость черепа, где вступают в обонятельную луковицу (лат. bulbus olfactorius) (здесь лежит тело второго нейрона), переходящую в обонятельный тракт (лат. tractus olfactorius), представляющий собой аксоны клеток, залегающих в (лат. bulbus olfactorius). Обонятельный тракт переходит в обонятельный треугольник (лат. trigonum olfactorium). Последний состоит преимущественно из нервных клеток и разделяется на две обонятельные полоски, вступающие в переднее продырявленное вещество (лат. substantia perforata anterior), лат. area subcallosa и прозрачную перегородку (лат. septum pellucidum), где находятся тела третьих нейронов. Затем волокна клеток этих образований различными путями достигают коркового конца обонятельного анализатора, залегающего в области крючка (лат. uncus) и парагиппокампальную извилины лат. gyrus parahyppocampalis височной доли больших полушарий мозга.
Обонятельные нервы — нервы специальной чувствительности.
Система обоняния начинается с обонятельной части слизистой носа (область верхнего носового хода и верхней части носовой перегородки). В ней находятся тела первых нейронов обонятельного анализатора. Эти клетки являются биполярными.
Как уже было отмечено выше, обонятельный анализатор представляет собой трёхнейронную цепь:Тела первых нейронов представлены биполярными клетками, находящимися в слизистой носа. Их дендриты оканчиваются на поверхности слизистой носа и образуют рецепторный аппарат обоняния. Аксоны этих клеток в виде обонятельных нитей заканчиваются на телах вторых нейронов, морфологически находящихся в обонятельных луковицах
Аксоны вторых нейронов формируют обонятельные тракты, которые оканчиваются на телах третьих нейронов в переднем продырявленном веществе (лат. substantia perforata anterior), лат. area subcallosa и прозрачной перегородке (лат. septum pellucidum)
Тела третьих нейронов также называются первичными обонятельными центрами. Важно отметить, что первичные обонятельные центры связаны с корковыми территориями как своей, так и противоположной стороны; переход части волокон на другую сторону происходит через переднюю спайку (лат. comissura anterior). Кроме этого, она обеспечивает связь с лимбической системой. Аксоны третьих нейронов направляются к передним отделам парагиппокампальной извилины, где расположено цитоархитектоническое поле Бродмана 28. В этой области коры представлены проекционные поля и ассоциативная зона обонятельной системы.
Запах, вызывающий аппетит, вызывает одновременно рефлекс слюноотделения, тогда как неприятный запах приводит к тошноте и рвоте. Эти реакции связаны с эмоциями. Запахи, могут быть приятными или неприятными. Основными волокнами, обеспечивающими связь обонятельной системы с автономными зонами мозга, являются волокна медиальных пучков переднего мозга и мозговых полосок зрительного бугра.
Медиальный пучок переднего мозга состоит из волокон, которые восходят от базальной обонятельной области, периминдалевидной области и ядер перегородки. На своём пути через гипоталамус часть волокон заканчивается на ядрах подбугорной области. Большинство же волокон направляется в ствол мозга и осуществляет контакт с вегетативными зонами ретикулярной формации, со слюноотделительными и дорсальными ядрами лат. n.intermedius (нерв Врисберга), языкоглоточного (лат. n. glossopharyngeus) и блуждающего (лат. n.vagus) нервов.
Мозговые полоски зрительного бугра отдают синапсы к ядрам поводка. От этих ядер к межножковому ядру (узлу Гансера) и к ядрам покрышки идёт поводково-ножковый путь, а от них волокна направляются к вегетативным центрам ретикулярной формации ствола мозга.
Волокна, осуществляющие связь обонятельной системы со зрительным бугром, гипоталамусом и лимбической системой, вероятно, и обеспечивают сопровождение обонятельных раздражителей эмоциями. Область перегородки, кроме прочих мозговых зон, связана через ассоциативные волокна с поясной извилиной (лат. gyrus cinguli).
30.Зрительный нерв, Типография, области инервации
Зрительный нерв представляет собой нерв специальной чувствительности. Зрительный нерв по своему развитию и строению представляет собой не типичный черепномозговой нерв, а как бы мозговое белое вещество, вынесенное на периферию и связанное с ядрами промежуточного мозга, а через них и с корой больших полушарий. Зрительный нерв берёт начало из ганглиозных клеток (третьих нервных клеток) сетчатки. Отростки этих клеток собираются в диске (или соске) зрительного нерва, находящемся на 3 мм ближе к середине от заднего полюса глаза. Далее пучки нервных волокон пронизывают склеру в области решётчатой пластинки, окружаются менингеальными структурами, образуя компактный нервный ствол. Нервные волокна изолированы друг от друга слоем миелина.
Среди пучков волокон зрительного нерва располагаются центральная артерия сетчатки (центральная ретинальная артерия) и одноимённая вена. Артерия возникает в центральной части глаза, а её капилляры покрывают всю поверхность сетчатки. Вместе с глазной артерией зрительный нерв проходит в полость черепа через зрительный канал, образованный малым крылом клиновидной кости.
Пройдя через толщу жирового тела глазницы зрительный нерв подходит к общему сухожильному кольцу. Этот его участок носит название лат. pars orbitalis. Затем он входит в зрительный канал (лат. canalis opticus) — эта часть носит название лат. pars intracanalicularis, а из глазницы в полость черепа выходит лат. pars intracranialis. Здесь в области предперекрёстной борозды клиновидной кости (лат. os sphenoidale) происходит частичный перекрёст волокон зрительного нерва — лат. chiasma opticum.
Латеральная часть волокон каждого из зрительных нервов направляется дальше по своей стороне.
Медиальная часть переходит на противоположную сторону, где соединяется с волокнами латеральной части зрительного нерва гомолатеральной (своей) стороны и образует вместе с ними зрительный тракт лат. tractus opticus.
По своему ходу ствол зрительного нерва окружён внутренним влагалищем зрительного нерва (лат. vagina interna n. optici), представляющим собой вырост мягкойоболочки головного мозга. Внутреннее влагалище щелевидным межвлагалищным пространством лат. spatia intervaginalis отделяется от наружного (лат. vagina externa n.optici), являющегося выростом паутинной и твёрдой оболочек головного мозга.
В лат. spatia intervaginalis проходят артерии и вены.
Каждый зрительный тракт огибает с боковой стороны ножку мозга (лат. pedunculus cerebri) и заканчивается в первичных подкорковых зрительных центрах, которые представлены с каждой стороны латеральным коленчатым телом, подушкой таламуса и ядрами верхнего холмика, где производится первичная переработка зрительной информации и формирование зрачковых реакций.
От подкорковых центров зрения нервы веером расходятся по обе стороны височной части головного мозга — начинается центральный зрительный путь (зрительная лучистость Грациоле), Далее волокна, несущие информацию от первичных подкорковых зрительных центров собираются вместе, чтобы пройти через внутреннюю капсулу. Заканчивается зрительный путь в коре затылочных долей (зрительной зоне) головного мозга.