

№1. Цикл лимонной кислоты – центральный процесс энергетического обмена.
Цикл Кребса (цикл трикарбоновых кислот, цикл лимонной кислоты) – циклический биохимический анаэробный процесс, в ходе которого освобождается водород и, окисляясь до воды, принимает участие в синтезе АТФ.
Главным источником низкомолекулярных предшественников для синтеза белков, жиров и углеводов является цикл цикл Кребса. Цикл Кребса — это центральный метаболический путь, характерный для всех клеток, дышащих кислородом. У эукариот цикл Кребса происходит внутри митохондрий, а у прокариот – в цитоплазме.
Этот важнейший цикл ферментативных реакций связывает процессы синтеза и распада. Цикл Кребса представляет собой первую стадию процессов анаболизма или третью стадию катаболизма.
Цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-КоА), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль «клеточного топлива»: углеводов, жирных кислот и аминокислот.
- - - - -
Дополнительно про сам цикл:
Основным материалом цикла служат восемь веществ, возникающих в клетке при многих других процессах: лимонная кислота, цис-аконитовая, изолимоиная, α-кетоглутаровая, янтарная, фумаровая, яблочная и щавелевоуксусная кислоты.
Продукты распада углеводов и жиров поступают в цикл Кребса в виде активированного остатка уксусной кислоты, которая образует тиоэфирную связь с коферментом А, а именно с его концевой БН-группой. Продукты распада белков поступают в цикл Кребса в виде ацетил-КоА, сукцинил-КоА, а-кетоглутаровой кислоты, а также янтарной и фумаровой кислот. Конечными продуктами одного оборота цикла Кребса, начиная со стадии включения ацетил-КоА, являются, две молекулы углекислого газа и четыре пары ионов водорода.
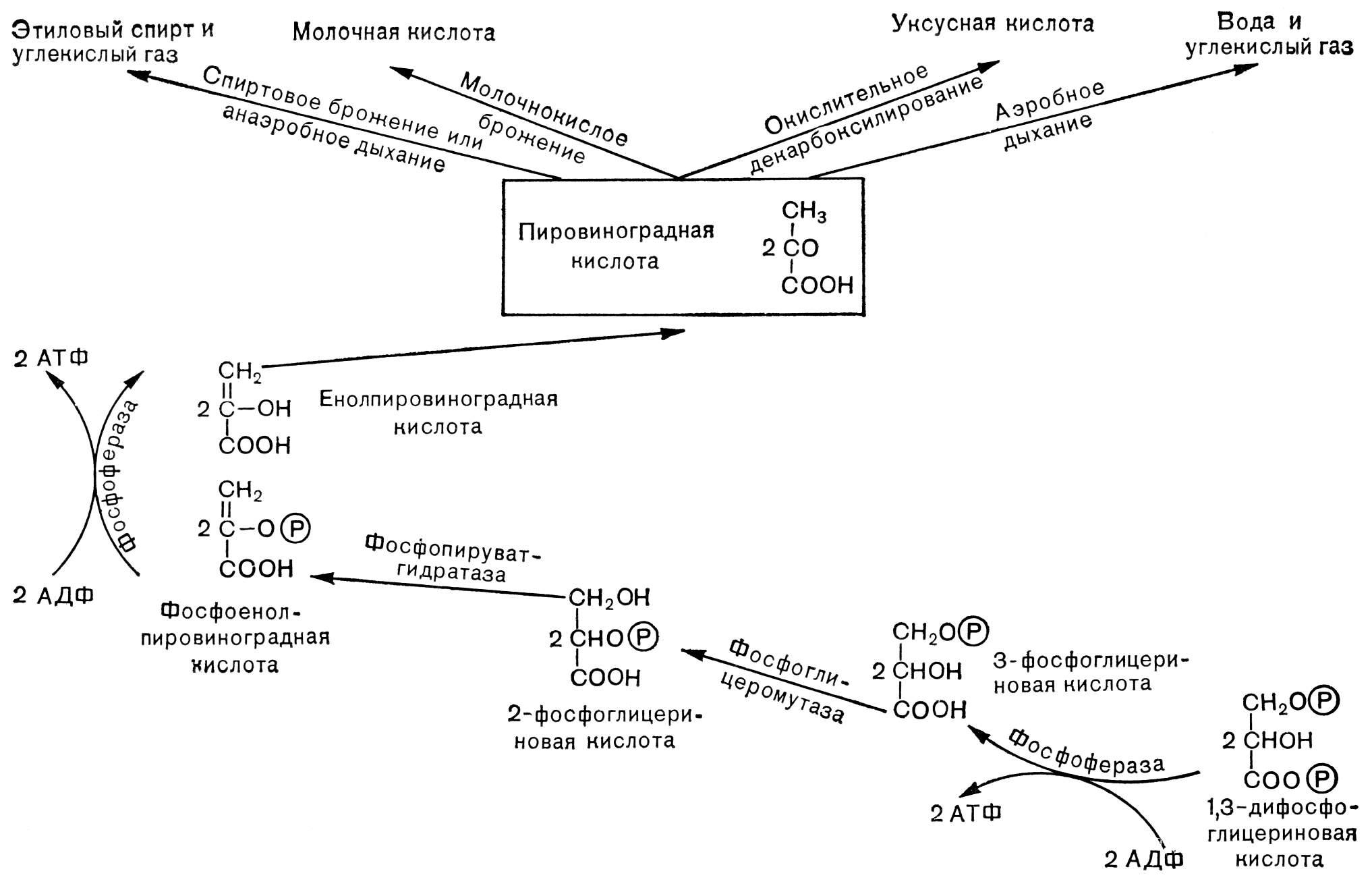
Цикл Кребса происходит в аэробных условиях и включает восемь стадий.
1. Конденсация ацетил-КоА с щавелевоуксусной кислотой, в результате чего образуется лимонная к-та, а кофермент А освобождается. Реакция катализируется цитрат-синтазой
2. Превращение лимонной к-ты в изолимонную при участии аконитат-гидратазы через промежуточную стадию цис-аконитата, связанного с ферментом.
3. Дегидрирование (так называется удаление из молекулы атомов водорода) цитрата с образованием a-кетоглутарата и СО, при участии изоцитратдегидрогеназы, которая функционирует при наличии Mg2+ и Мп2+.
4. Окислительное декарбоксилирование a-кетоглутарата до высокоэнергетического сукцинил-СоА. Реакция катализируется a-кетоглутаратдегидрогеназным комплексом (Mg2+).
5. Превращение сукцинил-СоА под влиянием сукцинил-СоА-синтетазы в сукцинат с отщеплением СоА.
6. Катализируемое сукцинатдегидрогеназой, содержащей ко-валентно связанный FAD и два железо-серных центра, дегидрирование сукцината с образованием фумарата.
7. Обратимая гидратация фумарата с образованием L-малата, катализируемая фумарат-гидратазой или фумаразой.
8. Катализируемое NAD-зависимой L-малатдегидрогеназой дегидрирование L-малата с образованием оксалоацетата. Эта реакция замыкает цикл Кребса и поставляет оксалоацетат для нового цикла. Большинство реакций цикла Кребса обратимы.
И чутка мнемоники:
Щуку ацетил лимонил,
Но нарцисса конь боялся,
Он над ним изолимонно
Альфа-кетоглутарался.
Сукцинился коэнзимом,
Янтарился фумарово,
Яблочек припас на зиму,
Обернулся щукой снова.
(щавелевоуксусная кислота, лимонная кислота, цис-аконитовая кислота, изолимонная кислота, α-кетоглутаровая кислота, сукцинил-KoA, янтарная кислота, фумаровая кислота, яблочная кислота, щавелевоуксусная кислота).
№2. Регулирование скорости ЦЛК.
Цикл Кребса подвержен регуляции; его скорость зависит от потребности в АТФ других метаболических реакций.
Регуляция происходит как на входе, так и в самом цикле.
Скорость цикла регулируется прежде всего за счет скорости образования «топлива» для этого цикла. Этим «топливом» является ацетил-КоА, образующийся при окислении пирувата и жирных кислот. Когда концентрация АТФ в митохондриях относительно велика и когда ацетил-КоА, а также промежуточные продукты цикла Кребса имеются в достаточном количестве, обеспечивающем удовлетворение энергетических нужд клетки, дальнейшее образование ацетил-КоА приостанавливается.
Рассмотрим теперь, каким образом регулируется сам цикл Кребса. Полагают, что общая скорость функционирования цикла Кребса определяется первой реакцией: Ацетил-КоА + Оксалоацетат → Цитрат + КоА.
Скорость цитрат-синтазной реакции регулируется концентрацией ее субстратов, в частности концентрацией ацетил-КоА, а она с свою очередь зависит от активности пируватдегидрогеназного комплекса. Регулируется эта реакция также концентрацией второго субстрата – оксалоацетата; возможно даже, что этот фактор играет главную роль, поскольку концентрация оксалоацетата в митохондриях очень низка и зависит от метаболитических условий. На активность цитрат-синтазы влияет также концентрация сукцинил-КоА, одного из более поздних промежуточных продуктов цикла.
№3. Пути образования активного ацетила.
… Не удалось найти что такое «активный ацетил», скорее всего имеется ввиду ацетил-Коэнзим А … Ацетил-КоА образуется во время второго шага кислородного клеточного дыхания, декарбоксилирования пирувата, который происходит в матриксе митохондрии. Ацетил-КоА затем поступает в цикл трикарбоновых кислот.
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
В окислении принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа (Е1), дигидролипоилацетилтрансфераза (Е2), дигидролипоилдегидрогеназа(Е3)) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД).
-На I стадии этого процесса окисления пируват теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента E1.
-На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом Е2.
-Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
**
- На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии фермента Е3 осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. --На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н+.
№4. Пути потребления активного ацетила.
Ацетил-КоА - макроэргический продукт конденсации коэнзима А с уксусной кислотой; в форме ацетил-КоА уксусная кислота участвует в различных обменных реакциях в организме, например в синтезе холестерина, стероидных гормонов, кетоновых тел.
При участии КоА осуществляются реакции цикла трикарбоновых кислот, окисления жирных к-т,, ацетилхолина. При многочисленных реакциях переносится не ацильная, а ацетильная rpуппа — остаток уксусной к-ты (ОССН3). Ацетил-КоА играет роль связующего звена между углеводным, жировым и белковым обменом.
№5. Назначение и пути потребления кетоновых тел.
АЦЕТОНОВЫЕ ТЕЛА (кетоновые тела) - группа продуктов обмена веществ, включающая b-оксимасляную кислоту, ацетоуксусную кислоту и ацетон. Образуются главным образом в печени и в меньшей степени в почках в результате неполного окисления жирных кислот, а также в процессе обмена углеводов и кетогенных аминокислот (фенилаланина, тирозина, лейцина и др.). Нормальное содержание кетоновых тел в крови КРС составляет 5—7 мг. Основня часть ацетоновых тел окисляется в цикле трикарбоновых кислот, частично они используются для синтеза стеринов, высших жирных кислот, фосфатидов, заменимых аминокислот. Небольшая часть А.т. выделяется из организма с мочой, молоком и выдыхаемым воздухом. При нарушении углеводно-липидпого обмена содержание А.т, в крови (ацетонемия) и моче (ацетанурия) может значительно повышаться, что приводит к развитию ацидоза и кетозов.
6.
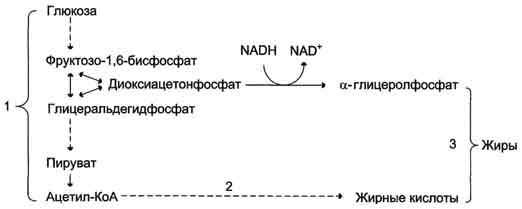
7. Синтез жира из углеводов.
1 - окисление глюкозы до пирувата и окислительное декарбоксилирование пирувата приводят к образованию ацетил-КоА; 2 - ацетил-КоА является строительным блоком для синтеза жирных кислот; 3 - жирные кислоты и а-глицеролфосфат, образующийся в реакции восстановления дигидроксиацетонфосфата, участвуют в синтезе триа-цилглицеролов.
8. Особенности превращения углеводов в пищеварительном тракте и в ходе метаболизма в организме жвачных.
Потребляемые углеводы представлены в основном дисахаридами(сахароза, лактоза) и полисахаридами (крахмал, гликоген). В ЖКТ подвергаются гидролитическому расщеплению L-( 1-4)-гликозидных связей . Целлюлозу могут переваривать только полигастричные животные, у них в рубце целюлозолитическая микрофлора. Расщипление начинается во рту под действием ферментов (амилаза,мальтаза,лактаза). Амилаза делает из крахмала мальтозу , часть которой расщепляется до 2 малекул глюкозы. Далее в желудке расщепляются под действием липаз и протеаз. Потом в тонком отделе кишечника гидролиз крахмала фермент (панкриотическая L-амилаза ).
В преджелудках взрослых жвачных, слипой кишке и толстом отделе киш-ке МОНОгастричных. Целлюлоза под действием целлюлазы гидролизуется до целлобиазы, которая потом под действием целлобиазы расщепляется до 2молекул глюкозы. Глюкоза распадается до летучих жирных кислот (уксусная, пропионовая, молочная), которые потом всасываются в кровь. Из крови в печень.
Вопрос 9. Биохимические механизмы поддержания нормального уровня глюкозы в крови.
в организме есть клетки, которые могут работать только на глюкозе - это нервные клетки и эритроциты. При быстром снижении уровня глюкозы в крови нервные клетки просто погибают. Это очень опасное явление известно под названием гипогликемия (дословно "низкое содержание глюкозы в крови"), и может привести к быстрой смерти. Без глюкозы мозг умирает всего за 10 секунд. Т.к. именно сахар в крови жизненно важен для жизни, то все силы организма направлены на поддержание в первую очередь уровня сахара в крови. При голодании, при отсутствии внешних поступлений пищи, организм должен поддерживать уровень сахара за счет своих ресурсов. Первым таким ресурсом являются запасы гликогена в печени и в мышцах. Этого источника питания хватает для поддержания уровня сахара в течение первых суток голодания. После первых суток голодания включается процесс получения глюкозы из тканей. В первую очередь это распад белков, вернее глюкогенных (из которых возможно получить глюкозу) аминокислот. Во вторую очередь - распад жиров на глицерин и жирные кислоты. Белки, которые активно распадаются начиная со второго дня голодания - это не что иное, как мышцы. Правильнее будет это назвать дистрофией мышц.
глюкагон повышает содержание глюкозы в крови:
-увеличивает мобилизацию гликогена;
-стимуляция глюконеогенеза – повышение работы ферментов.
адреналин вызывает гипергликемию:
-активируя мобилизацию гликогена;
глюкокортикоиды повышают глюкозу в крови:
-за счет подавление перехода глюкозы в клетку;
-стимуляция глюконеогенеза.