

4. Биосинтез первичных метаболитов и особенности регуляции по принципу обратной связи
4.1. Биосинтез первичных метаболитов
К первичным метаболитам относятся конечные продукты жизнедеятельности микроорганизмов, обладающие низкой молекулярной массой и используемые клеткой в качестве «строительных блоков» для жизненно важных макромолекул либо превращения в коферменты. Получаемые промежуточные продукты биосинтеза, которые отвечают указанному требованию, также рассматриваются как первичные метаболиты [5].
Другая группа первичных метаболитов включает полупродукты основных путей метаболизма – в частности, пентозофосфатного пути и цикла трикарбоновых кислот. Например, такие органические кислоты, как лимонная и фумаровая, относятся к первичным метаболитам. Продуцирование избыточного количества первичных метаболитов предотвращается большинством самих микроорганизмов, поскольку такой процесс истощает их и уменьшает способность к выживанию.
Микроорганизмы, способные к повышенному продуцированию, отбираются путем реализации специальных исследовательских программ по изысканию потенциальных «рабочих» культур для ферментации.
В ходе исследований с целью увеличения способности микроорганизмов к продуцированию нужных веществ и ингибирования регуляционных механизмов широко применяют генетические воздействия, а также изменяют условия окружающей среды.
4.2. Регуляция по принципу обратной связи
Регуляция по принципу обратной связи представляет собой метод контроля жизнедеятельности микроорганизмов для предотвращения чрезмерного синтеза аминокислот. Известны две методики [5], первая из которых основана на ограничении способности клетки к аккумуляции внутриклеточных ингибиторов и репрессоров, принадлежащих к конечным продуктам метаболизма, а вторая – на генетическом изменении самого фермента или системы, его образующей, до состояния, менее чувствительного к влиянию по принципу обратной связи. Для простой цепи превращений ограничение способности клетки к накоплению внутриклеточных ингибиторов или репрессоров часто применяют на практике с целью получения промежуточных продуктов реакций. Принцип такого ограничения показан на рис. 4.1. Если желательно аккумулировать в клетке промежуточный продукт С, сначала получают ауксотрофный (питательный) мутант, лишенный фермента С. Такой мутант требуется для роста Е. Если к клетке подводится недостаточное количество Е, организм не в состоянии аккумулировать ингибирующие или подавляющие концентрации Е, и тогда приводятся в действие реакции А и В, которые ингибируются регуляцией по принципу обратной связи. В клетке аккумулируются высокие концентрации С,
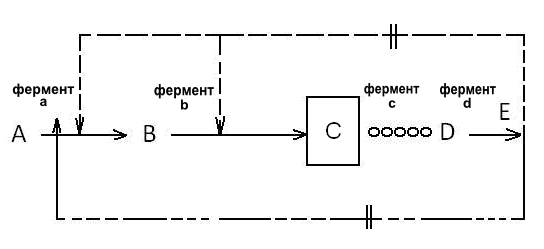
Рис. 4.1. Перепроизводство промежуточного продукта обмена
в простой цепи метаболических превращений
и этот продукт выделяется в среду. Это может быть использовано для получения надлежащих выходов определенных продуктов метаболизма и при разветвленных схемах биосинтетических превращений.
4.3. Накопление конечных продуктов метаболизма
Более сложной задачей является обеспечение преимущественного накопления конечных, а не промежуточных продуктов метаболизма. Она может быть решена при наличии разветвленной схемы биосинтетических превращений. Рассмотрим такую схему метаболизма, приводящую к получению конечных продуктов А и В. Часто удается интенсифицировать синтез продукта А путем уменьшения у клетке концентрации В. Примером использования подобного метода может служить ферментация лизина [5].
Лизин относится к аминокислотам, получаемым в промышленных масштабах ферментацией. Он важен как питательная добавка, поскольку большинство белков злаковых растений испытывают в нем дефицит. В бактериях лизин продуцируется по разветвленной схеме биосинтеза, в результате чего получают также метионин, треонин и изолейцин. После каждого разветвления первоначальные ферменты ингибируются соответствующими конечными продуктами и «перепроизводства» ферментов не происходит.
Однако организмы, применяемые при ферментации лизина (Corynebacterium glutamicum и Brevibacterium flavum), имеют, как показано на рис. 4.2, единственную аспартокиназу, получение которой регулируется путем согласованного ингибирования по принципу обратной связи треонином и лизином. При генетическом удалении гомосериндегидрогеназы образуется мутант, который не может расти, пока к среде добавляется метионин и
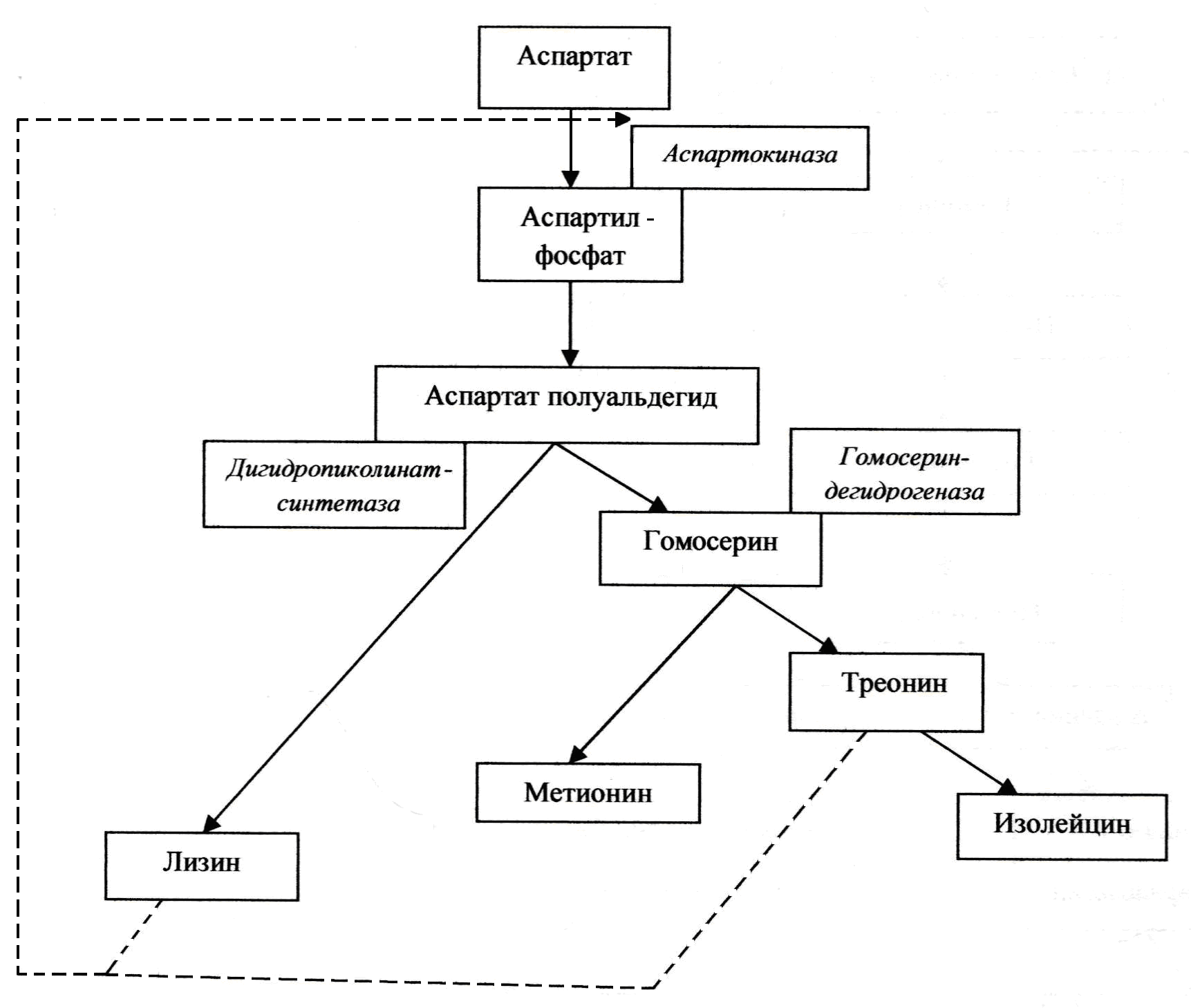
Рис. 4.2. Ферментация лизина
треонин. Пока добавки треонина в среду поддерживаются на низком уровне, внутриклеточная его концентрация является ограниченной и ингибирование аспартокиназы по принципу обратной связи байпассируется. По этой причине промежуточные продукты метаболизма отводятся в направлении лизинового ответвления схемы.
Можно отметить, что высокопродуктивные лизиновые штаммы отличаются следующими показателями:
– у них не наблюдается подавления аспартокиназы или аспартат-полуальдегиддегидрогеназы по принципу обратной связи;
– у них первый и второй ферменты лизинового ответвления не ингибируются и не подавляются лизином;
– у них отсутствует L-лизиндекарбоксилаза.
Такая специфичность позволяет мутантам продуцировать лизин в количестве не менее 50 г/л.