

1. 8 Классификация рефлексов. Классифицируя многообразные рефлексы организма человека и животного, учитывают их различные признаки и проявления.
Все рефлексы по происхождению делят на безусловные – врожденные – видовые рефлексы и условные – приобретенные в процессе индивидуальной жизни животного или человека, выработанные при определенных условиях.
На основании биологического значения тех или иных рефлексов для организма их делят на оборонительные, или защитные, направленные на удаление от раздражителя; пищевые, обеспечивающие добывание, потребление и переваривание пищи; половые, связанные с продолжением рода; ориентировочные, или исследовательские, проявляющиеся в повороте тела и движении в сторону нового раздражителя; позно-тонические, или рефлексы положения тела в пространстве, и локомоторные – перемещения тела в пространстве.
В зависимости от расположения рецепторов рефлекторной дуги, различают экстероцептивные рефлексы, возникающие в ответ на раздражение рецепторов поверхности тела, проприоцептивные – на раздражение рецепторов мышц, сухожилий и суставов, висцероцептивные – на раздражение рецепторов внутренних органов.
Классифицируют рефлексы и по органам, отвечающим на раздражение. При этом говорят о сердечных, дыхательных, сосудистых и других рефлексах.
Рефлексы различают также по характеру ответных реакций: секреторные, выражающиеся в выделении секрета, выработанного железой, трофические, связанные с изменением обмена веществ, и двигательные, или моторные, характеризующиеся сократительной деятельностью поперечнополосатых и гладких мышц. Последние составляют самую разнообразную группу рефлексов. При раздражении кожи возникают сгибательные, потирательные рефлексы, рефлексы почесывания и др. Раздражением кожи губ у ребенка вызывается сосательный рефлекс, роговицы глаза – мигательный. Сужение зрачка при действии света и его расширение в темноте представляют собой зрачковый рефлекс.
Двигательные проприоцептивные рефлексы возникают при раздражении рецепторов мышц и сухожилий. Так, при ударе по сухожилию четырехглавой мышцы бедра в результате ее растяжения возникает рефлекторное разгибание ноги в колене – коленный рефлекс, при ударе по ахиллову сухожилию – ахиллов рефлекс.
Рефлексы, заключающиеся в сужении и расширении сосудов, называются сосудодвигательными.
Двигательные рефлексы, возникающие при раздражении рецепторов гладкой мускулатуры внутренних органов, носят название висцеромоторных. С ними связаны движения желудка, кишечника, мочевого пузыря, мочеточников и др.
Все описанные выше рефлексы в зависимости от того, какие отделы центральной нервной системы участвуют в их осуществлении, делят на спинальные (осуществляющиеся с участием нейронов спинного мозга), бульбарные (с участием нейронов продолговатого мозга), мезенцефальные (с участием среднего мозга), диэнцефальные (с участием промежуточного мозга) и кортикальные (с участием нейронов коры больших полушарий головного мозга).
Примером спинальных рефлексов могут быть сгибательный, возникающий при пощипывании лапки лягушки пинцетом, потирательный – при раздражении кожи лягушки бумажкой, смоченной серной кислотой, и др. К ним же относятся рефлексы сухожильного типа.
Сосательный, мигательный рефлексы осуществляются с участием продолговатого, зрачковый – среднего мозга. Регуляция любой функции осуществляется с участием разных отделов центральной нервной системы. Поэтому классификация рефлексов по отделам мозга, участвующим в их осуществлении, относительна. Речь идет только о преимущественном значении нейронов того или иного отдела центральной нервной системы.
1. 9 Классификация и строение рецепторов. Живые организмы не могут существовать, не получая информации из внешней и внутренней среды. Восприятие энергии раздражителя и преобразование ее в нервный импульс осуществляют рецепторы.
Рецептор представляет собой окончание дендрита афферентного аксона или комплекс нервной и эпителиальной клеток, специально приспособленный для восприятия определенного типа раздражителей. Рецепторы являются начальными звеньями любой рефлекторной дуги, а также участвуют в оценке параметров полезного приспособительного результата в функциональных системах организма.
Дендрит каждого афферентного нейрона выходит на периферию и разветвляется на многочисленные тончайшие веточки – терминали. На конце каждой терминали находится утолщение – рецепторная бляшка.
По строению рецепторы подразделяют на первичные (первичночувствующие) и вторичные (вторичночувствующие) (рис. 10).
К первичным относят такие рецепторы, у которых действие раздражителя воспринимается непосредственно рецепторной бляшкой, которая может быть либо свободной, т. е. не иметь дополнительных образований, либо инкапсулированной. В этом случае рецепторная бляшка заключена в особую капсулу, осуществляющую первичное преобразование энергии раздражителя. К первичночувствующим рецепторам относятся: обонятельные, тактильные рецепторы и мышечные веретена.
К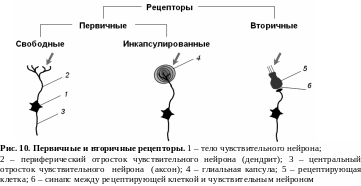 вторичным относят такие рецепторы, у
которых действие раздражителя
воспринимается специализированной
рецептирующей клеткой не нервного
происхождения. Возбуждение, возникшее
в рецептирующей клетке, передается
через синапс на чувствительный нейрон,
который возбуждается не непосредственно
стимулом, а опосредовано (вторично)
потенциалом рецептирующей клетки. К
вторичночувствующим рецепторам
относятся: рецепторы слуха, зрения,
вкуса, вестибулярные рецепторы.
вторичным относят такие рецепторы, у
которых действие раздражителя
воспринимается специализированной
рецептирующей клеткой не нервного
происхождения. Возбуждение, возникшее
в рецептирующей клетке, передается
через синапс на чувствительный нейрон,
который возбуждается не непосредственно
стимулом, а опосредовано (вторично)
потенциалом рецептирующей клетки. К
вторичночувствующим рецепторам
относятся: рецепторы слуха, зрения,
вкуса, вестибулярные рецепторы.
По расположению рецепторы подразделяют на: экстерорецепторы – воспринимают раздражители из внешней среды организма; интерорецепторы – воспринимают раздражители из внутренней среды организма. Они в свою очередь подразделяются на проприорецепторы – специализированные рецепторы опорно-двигательной системы (мышц, сухожилий, суставов), вестибулорецепторы – воспринимают положение тела в пространстве и висцерорецепторы – получают сигналы от внутренних органов.
По разнообразию воспринимаемых раздражителей рецепторы подразделяют на: мономодальные – приспособленные для восприятия только одного вида раздражителя, и полимодальные – приспособленные для восприятия различных раздражителей.
По модальности рецепторы подразделяют на: хеморецепторы – воспринимают действие химических веществ; фоторецепторы – воспринимают световые раздражители; механорецепторы – воспринимают давление, вибрацию, перемещение, степень растяжения; терморецепторы – чувствительны к изменениям температуры; ноцицепторы – воспринимают болевое раздражение.
По способности воспринимать раздражитель, находящийся на расстоянии от рецептора или при непосредственном контакте с ним, различают дискантные и контактные рецепторы
Все рецепторы обладают следующими свойствами.
1. Специфичность – способность воспринимать определенный, т. е. адекватный данному рецептору, раздражитель. Эта способность рецепторов сформировалась в процессе эволюции.
2. Высокая чувствительность – способность реагировать на очень малые по интенсивности параметры адекватного раздражителя. Например, для возбуждения фоторецепторов сетчатки глаза достаточно нескольких, а иногда и одного, квантов света. Обонятельные рецепторы информируют организм о появлении в атмосфере единичных молекул пахучих веществ.
3. Способность к ритмической генерации импульсов возбуждения в ответ на однократное действие раздражителя.
4. Возбуждение рецепторов наступает после некоторого латентного периода и имеет последействие.
5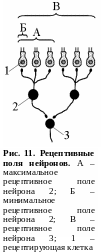 .
Способность к адаптации – т. е. способность
приспосабливаться ("привыкать") к
постоянно действующему стимулу. Адаптация
может выражаться в снижении активности
рецептора и частоты генерации импульсов
возбуждения, вплоть до полного его
прекращения. В зависимости от скорости
адаптации различают: быстроадаптирующиеся
(тактильные), медленноадаптирующиеся
(терморецепторы), неадаптирующиеся
(болевые, вестибулярные и проприорецепторы).
.
Способность к адаптации – т. е. способность
приспосабливаться ("привыкать") к
постоянно действующему стимулу. Адаптация
может выражаться в снижении активности
рецептора и частоты генерации импульсов
возбуждения, вплоть до полного его
прекращения. В зависимости от скорости
адаптации различают: быстроадаптирующиеся
(тактильные), медленноадаптирующиеся
(терморецепторы), неадаптирующиеся
(болевые, вестибулярные и проприорецепторы).
Выделяют несколько видов адаптации: изменение возбудимости рецептора в сторону снижения – десенсибилизация; изменение возбудимости в сторону повышения – сенсибилизация.
При действии сильных раздражителей, порог чувствительности повышается. Например: если человек вышел из тепла сразу в холод – то человеку вначале холодно, но затем рецепторы повышают свой порог и человеку становится тепло.
Р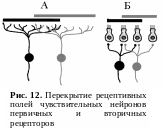 ецептивное
поле. Рецептивным полем нейрона называют
множество рецепторов, функционально
связанных с этим нейроном. Рецептивное
поле нейрона представляет собой
динамическое образование – один и тот
же нейрон в различные моменты времени
может оказаться функционально связанным
с различным количеством рецепторов.
Максимальная величина рецептивного
поля какого-либо нейрона соответствует
количеству рецепторов, которые связаны
с эти нейроном морфологически, а
минимальная величина может ограничиваться
всего одним рецептором (рис. 11).
ецептивное
поле. Рецептивным полем нейрона называют
множество рецепторов, функционально
связанных с этим нейроном. Рецептивное
поле нейрона представляет собой
динамическое образование – один и тот
же нейрон в различные моменты времени
может оказаться функционально связанным
с различным количеством рецепторов.
Максимальная величина рецептивного
поля какого-либо нейрона соответствует
количеству рецепторов, которые связаны
с эти нейроном морфологически, а
минимальная величина может ограничиваться
всего одним рецептором (рис. 11).
У первичных рецепторов зоны ветвления периферических отростков чувствительных нейронов могут перекрывать друг друга (рис. 12, А). У вторичных рецепторов одна рецептирующая клетка может контактировать с несколькими чувствительными нейронами, т. е. может входить в состав рецептивных полей различных нейронов (рис. 12, Б).
Рецептивным полем рефлекса называют совокупность рецепторов, раздражение которых вызывает данный тип рефлекса.
1. 10 Понятие о нервном центре. Свойства нервных центров. В сложных многоклеточных организмах животных и человека отдельная нервная клетка не в состоянии регулировать какие-либо функции. Все основные формы деятельности нервной системы связаны с участием в ее функциях определенных групп нервных клеток – нервных центров. Нервный центр – центральный (третий) компонент рефлекторной дуги, где происходит переработка информации, вырабатывается программа действия, формируется эталон результата.
Нервным центром называется совокупность нейронов, расположенных на различных уровнях ЦНС и регулирующих сложный рефлекторный процесс или функцию. В нервном центре выделяют следующие отделы: низший или исполнительный, рабочий и высший или регуляторный.
Низший (исполнительный) отдел нервного центра локализован в спинном мозге и передает информацию от рабочего отдела к рабочим органам.
Рабочий отдел нервного центра – это отдел, ответственный за осуществление данной функции, как правило, находится в мозговом стволе.
Высший (регуляторный) отдел нервного центра расположен в коре больших полушарий мозга и регулирует активность рабочего отдела нервного центра; вмешивается в регуляцию функций эпизодически, т. е. в тех случаях, когда необходимо внести поправку в автоматическую деятельность рабочего отдела. Высшие отделы вмешиваются в работу нервного центра по условно-рефлекторному механизму.
Активность регуляторного (высшего) отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию и от внешних стимулов среды.
Вышеизложенное можно рассмотреть на примере дыхательного центра. Исполнительный (низший) отдел дыхательного нервного центра расположен в передних рогах грудного отдела спинного мозга и транслирует приказы рабочего центра к дыхательным мышцам. Рабочий отдел представлен центрами вдоха, выдоха и пневмотаксиса, расположенными в продолговатом мозге и варолиевом мосту. Нарушение этого отдела вызывает остановку дыхания. Регуляторный (высший) отдел дыхательного центра расположен в лобной доле коры больших полушарий и позволяет произвольно регулировать легочную вентиляцию (глубину и частоту дыхания). Однако эта произвольная регуляция небезгранична и зависит от функциональной активности рабочего отдела, афферентной импульсации, отражающей состояние внутренней среды (в данном случае рН крови, концентрации СО2 и О2 в крови).
Свойства нервных центров. Физиологические свойства нервных центров определяют характер ответных реакций. Эти свойства в значительной мере связаны с особенностями проведения нервных импульсов через синапсы, соединяющие различные нервные клетки.
1. Одностороннее проведение возбуждения. Обусловлено особенностями проведения возбуждения по синапсу. Медиаторы выделяются в пресинаптическом окончании; рецепторы, взаимодействующие с медиаторами, находятся в постсинаптической мембране.
2. Замедление проведения возбуждения – наличие латентного периода (периода от начала действия сигнала до проявления рефлекторного акта). Обусловлено синаптической задержкой в центральной части рефлекторной дуги, где возбуждение проходит через синапсы. На выделение и диффузию медиатора в синапсе требуется промежуток времени в 1,5-2 мс (синаптическая задержка). Чем больше нейронов в рефлекторной дуге, тем продолжительнее время рефлекса.
3. Рефлекторный ответ зависти от силы и продолжительности действия раздражителя.
4. Суммация возбуждений. В работе нервных центров значительное место занимают процессы пространственной и временнóй суммации подпороговых раздражений. Временнáя суммация наблюдается, если множество слабых импульсов приходит к нейрону по одному и тому же пути через один синапс с коротким интервалом времени. В результате из пресинаптической мембраны выбрасывается множество квантов медиатора, происходит суммирование изменений на постсинаптической мембране, и она деполяризуется до уровня, достаточного для генерации возбуждения. Пространственная суммация связана с суммированием подпороговых потенциалов, возникающих одновременно в разных синапсах одного нейрона. Оба вида суммации происходят в области аксонного холмика, где и генерируется ПД.
5. Низкая лабильность обусловлена сравнительно низкой скоростью развития синаптической передачи нервного импульса в химическом синапсе (около 50-100 импульсов в секунду, что в 5 – 6 раз ниже лабильности аксона).
6. Утомление нервных центров. Длительное повторное раздражение рецептивного поля рефлекса приводит к ослаблению рефлекторной реакции вплоть до полного исчезновения, что называется утомлением. Этот процесс связан с деятельностью синапсов – в последних наступает истощение запасов медиатора, уменьшаются энергетические ресурсы, происходит адаптация постсинаптического рецептора к медиатору.
Разные нервные центры имеют различную скорость утомления. Менее утомляемы центры вегетативной нервной системы, координирующие работу внутренних органов. Значительно более утомляемы центры соматической нервной системы, управляющие произвольной скелетной мускулатурой.
7. Трансформация ритма. Сигналы, которые уходят от нервного центра по своим параметрам (частота, сила) отличаются от таковых пришедших к нервному центру. Нервные клетки обладают свойством изменять частоту передающихся импульсов, т. е. свойством трансформации ритма.
Характер ответного разряда нейрона зависит от свойств раздражителя, а также и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). В обычных условиях, чем сильнее раздражение, тем чаще проявляется ответная импульсация.
При высокой возбудимости нейрона (например, после приема кофеина) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы, наконец, достичь порога возникновения потенциала действия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции организма на внешние раздражения.
8. Нервные центры очень чувствительны к недостатку кислорода. В нервных клетках осуществляется интенсивный обмен веществ, для чего необходима энергия и постоянный приток нужного количества кислорода. Особенно чувствительны к недостатку кислорода нервные клетки коры больших полушарий головного мозга: после 5 – 6 минут кислородного голодания они погибают. У человека даже кратковременное ограничение мозгового кровообращения приводит к потере сознания. Недостаточное снабжение кислородом легче переносят нервные клетки мозгового ствола: их функция восстанавливается через 15 – 20 минут после полного прекращения кровоснабжения. А функция клеток спинного мозга восстанавливается даже после 30 минут отсутствия кровоснабжения.
9. Нервные центры чувствительны к химическим веществам, которые проходят через гематоэнцефалический барьер. Нервные центры обладают специфической реакцией на различные фармакологические вещества, что говорит о своеобразии протекающих в них процессов. Стрихнин возбуждает нервные центры, блокируя работу тормозных синапсов. Хлороформ и эфир сначала возбуждают, а затем подавляют работу нервных центров. Апоморфин возбуждает рвотный центр, цититон и лобелин – дыхательный центр, а морфин угнетает его работу. Коразол возбуждает клетки двигательной зоны коры, вызывая эпилептические судороги.
10. Нервные центры всегда находятся в тонусе. Это обусловлено тем, что они постоянно получают импульсацию от других структур мозга. Кроме того, к нервным центрам постоянно идут импульсы обратной афферентации от рецепторов исполнительных органов. В ответ центры посылают редкие импульсы к органам, поддерживая в них соответствующий тонус. Даже во время сна мышцы не расслабляются полностью и контролируются соответствующими центрами.
11. Последействие нервных центров. Следовые процессы или последействие означает, что после окончания действия раздражителя активное состояние нервного центра продолжается еще некоторое время. Длительность следовых процессов различна. В спинном мозге – несколько секунд или минут. В подкорковых центрах мозга – десятки минут, часы и даже дни. В коре больших полушарий – до нескольких десятков лет.
Следовые процессы имеют важное значение в понимании механизмов памяти. Непродолжительное последействие до 1 часа связано с циркуляцией (реверберацией) импульсов в нервных цепях (Р. Лоренте де Но, 1934) и обеспечивает кратковременную память. Механизмы долговременной памяти основаны на изменении структуры белков. В процессе запоминания, согласно биохимической теории памяти (Х. Хиден, 1969) происходят структурные изменения в молекулах РНК, на основе которых строятся измененные белки с отпечатками прежних раздражителей. Эти белки длительно содержатся в нейронах, а также в глиальных клетках головного мозга.
1. 11 Торможение в ЦНС. Торможение в ЦНС– это активный процесс, проявляющийся в подавлении или ослаблении возбуждения. В отличие от возбуждения, торможение по нервным волокнам не распространяется.
Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г. Значительно позже английский физиолог Шеррингтон открыл, что процессы возбуждения и торможения участвуют в любом рефлекторном акте.
Значение торможения:
1) Координационное – процесс торможения обеспечивает упорядоченность или координацию в работе нервных центров, например, чтобы согнуть руку, надо возбудить центр сгибания, посылающий нервные импульсы на бицепс, и затормозить центр разгибания, посылающий нервные импульсы на трицепс.
2) Охранительное значение – при действии сверхсильных раздражителей в нервном центре развивается не возбуждение, а торможение, в результате восстанавливаются запас АТФ и медиатора.
3) Ограничение притока афферентных импульсов в ЦНС (так как мозг не в состоянии переработать весь объем второстепенной малозначимой для человека информации).
Важнейшая роль торможения в координационной деятельности ЦНС наглядно прослеживается при введении в организм подопытного животного определённой дозы стрихнина, временно лишающего ЦНС способности развивать торможение. У такого животного практически исчезает всякая координация рефлексов: любое локальное раздражение вызывает одновременную двигательную реакцию всей мускулатуры тела.
По месту возникновения различают пресинаптическое торможение и постсинаптическое. При пресинаптическом торможении тормозной эффект реализуется на пресинаптической мембране, этот вид торможения участвует в ограничении притока чувствительных импульсов в мозг. Возникает на аксо-аксональных синапсах.
Постсинаптическое торможение осуществляется на постсинаптической мембране. Это основной вид торможения. Развивается в истинных тормозных синапсах (аксосоматических и аксодендрических) с участием тормозных медиаторов, которые изменяют свойства постсинаптической мембраны таким образом, что способность нервной клетки генерировать процессы возбуждения подавляется.
По нейронной организации торможение подразделяют на поступательное, возвратное, латеральное (боковое) и реципрокное.
1. Поступательное торможение обусловлено включением тормозных нейронов на пути следования возбуждения.
2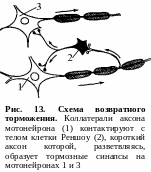 .
Возвратное торможение осуществляется
вставочными тормозными нейронами
(клетками Реншоу). Импульсы от мотонейронов,
через отходящие от его аксона коллатерали,
активируют клетку Реншоу, которая в
свою очередь вызывает торможение
разрядов данного мотонейрона (рис. 13).
Это торможение реализуется за счет
тормозных синапсов, образованных клеткой
Реншоу на теле активирующего ее
мотонейрона. Таким образом, из двух
нейронов формируется контур с отрицательной
обратной связью, которая дает возможность
стабилизировать частоту разряда
мотонейрона и подавлять избыточную его
активность.
.
Возвратное торможение осуществляется
вставочными тормозными нейронами
(клетками Реншоу). Импульсы от мотонейронов,
через отходящие от его аксона коллатерали,
активируют клетку Реншоу, которая в
свою очередь вызывает торможение
разрядов данного мотонейрона (рис. 13).
Это торможение реализуется за счет
тормозных синапсов, образованных клеткой
Реншоу на теле активирующего ее
мотонейрона. Таким образом, из двух
нейронов формируется контур с отрицательной
обратной связью, которая дает возможность
стабилизировать частоту разряда
мотонейрона и подавлять избыточную его
активность.
3. Латеральное торможение, т. е. возникает процесс торможения группы нейронов, расположенных рядом с группой возбужденных клеток. Этот вид торможения распространен в сенсорных системах.
4. Реципрокное торможение, т. е. процесс, основанный на том, что сигналы по одним и тем же афферентным путям обеспечивают возбуждение одной группы нейронов, а через вставочные тормозные клетки вызывают торможение другой группы нейронов. Этот тип торможения называют также сопряженным. Проявляется, например, на уровне мотонейронов спинного мозга, иннервирующих мышцы-антагонисты (сгибатели – разгибатели конечностей). При сгибании руки или ноги центры мышц-разгибателей затормаживаются. Рефлекторный акт возможен только при сопряженном торможении мышц-антагонистов. При ходьбе сгибание ноги сопровождается расслаблением разгибателей и, наоборот, при разгибании тормозятся мышцы-сгибатели. Если бы этого не происходило, то возникла бы механическая борьба мышц, судороги, а не приспособительные двигательные акты.
В процессе онтогенеза за счет развития тормозных нейронов формируются тормозные механизмы ЦНС. Ранней их формой является постсинаптическое торможение, позднее формируется пресинаптическое торможение. Благодаря формированию тормозных механизмов существенно ограничивается иррадиация возбуждения в ЦНС, свойственная новорожденным, появляются индукционные отношения. Безусловные рефлексы становятся более точными и локализованными.
1. 12 Координация рефлекторной деятельности. Координация рефлексов – это согласованное взаимодействие нервных центров для обеспечения какого-либо процесса. Каждый рефлекс представляет собой реакцию всей ЦНС и зависит от всей совокупности межцентральных взаимоотношений. Координация функций обеспечивает рефлекторные акты, соответствующие воздействиям внешней среды. Эти рефлекторные акты могут проявляться со стороны различных систем (мышечной, эндокринной, сердечно-сосудистой). К примеру: при беге рефлекторно работают мышцы сгибателей и разгибателей. Повышается артериальное давление, увеличивается просвет сосудов, учащается биение сердца и дыхания. Все это скоординировано. Координация функций определяется характеристикой взаимоотношений между рефлекторными проявлениями со стороны различных систем организма для реализации определённого физиологического акта. Координационные механизмы достигают своего совершенства к 18 – 20 годам.
Основой координации служат следующие особенности ЦНС.
1. Конвергенция – концентрация импульсов на 1 вставочном нейроне. В ЦНС большинство нейронов получают информацию от разных источников благодаря конвергенции. Это обеспечивает пространственную суммацию импульсов и усиление конечного эффекта. Конвергенция на клетках нервного центра разных афферентных входов предопределяет важные интегративные, перерабатывающие информацию функции центральных нейронов, т. е. высокий уровень интеграционных функций.
2. Дивергенция. Способность нейрона устанавливать многочисленные синаптические связи с различными нервными клетками в пределах одного или разных нервных центров называется дивергенцией. Например, центральные окончания аксонов первичного афферентного нейрона образуют синапсы на многих вставочных нейронах. Благодаря этому одна и та же нервная клетка может участвовать в различных нервных реакциях и контролировать большое число других нейронов, что приводит к иррадиации возбуждения.
3. Наличие в ЦНС двух типов вставочных нейронов: возбуждающих и тормозных. Постоянное взаимодействие процессов возбуждения и торможения, как между разными центрами, так и в пределах каждого из них является важнейшей характеристикой их деятельности.
Механизмы координации
1. Иррадиация возбуждения. Механизм иррадиации базируется на дивергенции терминалей афферентных и вставочных нейронов. Это самый древний механизм координации.
Нейроны разных центров связаны между собой многочисленными вставочными нейронами, поэтому при раздражении рецепторов возбуждение может распространяться не только на нейроны центра данного рефлекса, но и на другие нейроны. Распространение процессов возбуждения на другие нервные центры называют явлением иррадиации. Чем сильнее и длительнее афферентное раздражение и чем выше возбудимость окружающих нейронов, тем больше нейронов охватывает процесс иррадиации. Процессы торможения ограничивают иррадиацию и способствуют концентрации возбуждения в исходном пункте ЦНС.
Процесс иррадиации играет важную положительную роль при формировании новых реакций организма (ориентировочных реакций, условных рефлексов). Благодаря иррадиации возбуждения между различными нервными центрами возникают новые функциональные связи – условные рефлексы. Иррадиация возбуждения может оказать отрицательное воздействие на состояние и поведение организма, нарушая тонкие взаимоотношения между возбужденными и заторможенными нервными центрами и вызывая нарушения координации движений.
2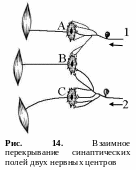 .
Облегчение и окклюзия. Конвергенция
множества путей, т. е. нервных цепочек
на одной группе мотонейронов лежит в
основе феноменов облегчения и окклюзии.
Облегчение и окклюзия возможны в том
случае, когда на какой-то части нейронов
сходятся (конвергируют) пресинаптические
волокна из различных афферентных
источников (рис. 14).
.
Облегчение и окклюзия. Конвергенция
множества путей, т. е. нервных цепочек
на одной группе мотонейронов лежит в
основе феноменов облегчения и окклюзии.
Облегчение и окклюзия возможны в том
случае, когда на какой-то части нейронов
сходятся (конвергируют) пресинаптические
волокна из различных афферентных
источников (рис. 14).
Облегчение – это превышение эффекта одновременного действия двух слабых пороговых раздражителей над суммой их раздельных эффектов. Например, нейроны А и С получают от каждого афферентного нейрона (1 и 2) достаточное количество синаптических окончаний (например, по 4), чтобы в них возникло возбуждение (рис. 14). Нейрон В получает от тех же афферентных нейронов (1 и 2) меньшее число окончаний (по 2), поэтому их импульсов будет недостаточно, чтобы генерировать в этом нейроне нервный импульс, а возникает лишь подпороговое возбуждение. Вследствие этого, при раздельном раздражении афферентных нейронов 1 и 2 возникают рефлекторные реакции только в нейронах А и С. Но при одновременном раздражении афферентных нейронов 1 и 2 нервный импульс генерируется и нейроном В (происходит суммация нервных импульсов от двух источников). Поэтому выраженность такого суммарного рефлекторного ответа будет больше. Это явление получило название центрального облегчения.
Окклюзия (закупорка) – это явление, противоположное облегчению. Окклюзия возникает при действии сильных надпороговых раздражителей. При взаимном перекрывании синаптических полей двух нервных центров суммарный результат оказывается меньшим, чем сумма отдельных реакций. Так, при раздражении афферентного поля первого нервного центра возникает возбуждение в двух нейронах (А и В), при активации второго центра также возникает возбуждение в двух центрах (С и В), однако при одновременной стимуляции обоих центров (1 и 2) суммарный двигательный ответ равен только трем единицам (возбуждаются центры А, В и С), а не четырем (рис. 14). Это обусловлено тем, что один и тот же мотонейрон (В) относится одновременно к обоим нервным центрам. Явление окклюзии приводит к снижению силы суммарной ответной реакции.
3. Принцип общего конечного пути. Афферентных нейронов в центральной нервной системе в несколько раз больше, чем эфферентных. В связи с этим многие афферентные влияния поступают к одним и тем же вставочным и эфферентным нейронам, которые являются для них общими конечными путями к рабочим органам. Система реагирующих нейронов образует, таким образом, как бы воронку («воронка Шеррингтона»). Множество разнообразных раздражений может возбудить одни и те же мотонейроны спинного мозга и вызвать одну и ту же двигательную реакцию. Так, например, мотонейроны, иннервирующие дыхательную мускулатуру, помимо обеспечения вдоха участвуют в таких рефлекторных реакциях, как чихание, кашель и др.
Английский физиолог Ч. Шеррингтон, установивший принцип общего конечного пути, предложил различать союзные и антагонистические рефлексы. Встречаясь на общих конечных путях, союзные рефлексы взаимно усиливают друг друга, а антагонистические – тормозят. В первом случае в нейронах общего конечного пути имеет место пространственная суммация (например, сгибательный рефлекс усиливается при одновременном раздражении нескольких участков кожи). Во втором случае происходит борьба конкурирующих влияний за обладание общим конечным путем, в результате чего один рефлекс осуществляется, а другие затормаживаются. При этом освоенные движения выполняются с меньшим трудом, так как в их основе лежат упорядоченные во времени синхронизированные потоки импульсов, которые проходят через конечные пути легче, чем импульсы, поступающие в случайном порядке.
Преобладание на конечных путях той или иной рефлекторной реакции обусловлено ее значением для жизнедеятельности организма в данный момент.
В таком отборе важную роль играет наличие в центральной нервной системе доминанты. Она обеспечивает протекание главной реакции. Например, ритмический шагательный рефлекс и одиночный, непрерывный рефлекс сгибания при болевом раздражении являются антагонистическими. Однако спортсмен, внезапно получивший травму, может продолжать бег к финишу, т. е. осуществлять ритмический рефлекс и подавлять болевые раздражения, которые, поступая к мотонейронам сгибательных мышц, препятствуют попеременному сгибанию и разгибанию ноги.
4. Обратная связь или вторичная афферентация. Всякий двигательный акт, вызванный афферентным раздражителем, сопровождается возбуждением рецепторов мышц, сухожилий, суставных сумок. Сигналы с проприорецепторов вторично поступают в ЦНС, что позволяет осуществлять коррекцию ее деятельности и саморегуляцию в соответствии с текущими потребностями организма и наличной обстановкой. Этот важный принцип рефлекторной саморегуляции функций организма называется принципом обратной связи.
Кроме того, за счет обратной связи поддерживается тонус нервных центров.
5. Реципрокные (сопряженные) взаимоотношения между нервными центрами. В основе взаимосвязи между нервными центрами лежит процесс индукции – развитие (индуцирование) противоположного процесса. Индукция ограничивает распространение (иррадиацию) нервных процессов и обеспечивает концентрацию.
Различают одновременную и последовательную индукцию. Сильный процесс возбуждения в нервном центре вызывает (индуцирует) торможение в соседних нервных центрах (одновременная отрицательная индукция), а сильный тормозной процесс индуцирует в соседних нервных центрах возбуждение (одновременная положительная индукция). Так, для сгибательного движения в суставе необходимо не только сокращение мышц-сгибателей, но и одновременное расслабление мышц- разгибателей. При этом в мотонейронах мышц-сгибателей возникает процесс возбуждения, а в мотонейронах мышц-разгибателей – процесс торможения. При возбуждении же центров разгибателей, наоборот, тормозятся центры сгибателей.
При смене процессов возбуждения и торможения в пределах одного центра говорят о последовательной отрицательной или положительной индукции. Она имеет большое значение при организации ритмической двигательной деятельности, обеспечивая попеременное сокращение и расслабление мышц.
Установлено, что у человека во время ходьбы и бега основной формой координации являются реципрокные отношения, но помимо них имеются фазы одновременной активности мышц-антагонистов голеностопного и особенно коленного и тазобедренного суставов. Длительность фаз одновременной активности увеличивается с повышением скорости перемещения.
У детей четкие индукционные взаимоотношения между процессами торможения и возбуждения начинают развиваться в возрасте с 3-х до 5-и лет, так как в этом возрасте возрастает сила нервных процессов.
6. Доминанта. Активность нервных центров непостоянна, и преобладание активности одних из них над активностью других вызывает заметные перестройки в процессах координации рефлекторных реакций. В 1923 г. А. А. Ухтомский сформулировал принцип доминанты как рабочий принцип деятельности нервных центров.
Доминанта – это временное преобладание одного нервного центра или группы центров над другими, определяющее текущую деятельность организма.
Основные черты, доминанты следующие: 1) повышенная возбудимость нервных центров; 2) стойкость возбуждения во времени; 3) способность усиливать свое возбуждение за счет суммации нервных импульсов, идущих в другие центры (в результате этого раздражение различных рецепторных полей начинает вызывать рефлекторный ответ, характерный для деятельности этого доминантного центра); 4) доминирующий центр по механизму одновременной индукции вытормаживает другие центры.
Доминантный очаг в ЦНС может возникать под влиянием разных факторов, в частности, сильной афферентной стимуляции, гормональных воздействий, изменения химизма крови, мотиваций и т. д.
Несмотря на относительную стойкость и инертность возбуждения в доминантном очаге, деятельность ЦНС в нормальных условиях существования весьма динамична и изменчива. ЦНС обладает способностью к перестройке доминантных отношений в соответствии с изменяющимися потребностями организма, и в течение всей жизни человека одна доминанта сменяет другую. Сильные доминанты остаются даже во сне.
Доминантный очаг у ребенка возникает быстрее и легче, чем у взрослых, но для него характерна низкая устойчивость к внешним раздражителям. С этим в значительной степени связана неустойчивость внимания у детей: новые раздражители легко вызывают новую доминанту.
7. Пластичность нервных центров. Под пластичностью понимают функциональную изменчивость и приспособляемость нервных центров, их способность выполнять новые, необычные для них рефлекторные акты. Это особо ярко проявляется после удаления различных отделов мозга. Если были частично удалены какие-то отделы мозжечка или коры больших полушарий, нарушенная функция со временем может восстановиться.
Г лава
2. ЧАСТНАЯ ФИЗИОЛОГИЯ
лава
2. ЧАСТНАЯ ФИЗИОЛОГИЯ
ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ