

4.2. Система голонасінних
Дендрологи користуються однією з двох, наведених в табл. 7.1, класифікацій деревних рослин: або за А.Енглером, або за А.Л. Тахтаджяном. Хід еволюції насінних рослин представлений на рис.1.
Виділення А.Енглером і його послідовниками двох підвідділів голонасінних Cycadophytina і Coniferophytina зумовлюються наявними двома лініями еволюції, які на думку авторів (наприклад, А.Швейковська і Є.Швейковський, 1986), мали спільних предків, а надалі незалежно одна від другої досягли одинакових рівнів організації. Перша еволюційна лінія (рис. 7.2) розвивалася із збереженням відносно багатьох рис папоротеподібних: великі листки, дуже почленована пластинка; слабкий приріст стебла в товщину, його будова маноксильна, стебло малогалузисте або негалузисте.
Еволюція тут йшла в напрямках спрощення будови листка до цілісної пластинки у роду Gnetum, скорочення спороносних листків і обмеження розмірів спеціалізованих структур - стробілів. Спочатку стробіли були двостатеві (Bennetitopsida), надалі одностатевими. Деякі особливості цієї групи голонасінних зближають їх з покритонасінними рослинами: наявність судин у вторинній ксилемі гнетових, двостатеві стробіли беннетитових, редукція чоловічого гаметофіту до одноклітинної ( але багатоядерної) структури, редукція архегоніїв у гнетових. Саме тут, зазначають згадані автори, серед крупнолистих голонасінних треба шукати ппепкові сіюоми покоитонасінних рослин.
Ери |
Палеозойська |
Мезозойська |
Кайнозойська |
|||||
Періоди (млн. років назад) |
Девонський, 408 (девон) |
Кам’яновугільний, 360 (карбон) |
Пермський, 286 (перм) |
Тріасовий, 248 (тріас) |
Юрський, 213 |
Крейдяний, 144 (крейда) |
Третинний, 65 |
Четвертинний, 2 |

Рис.1. Хід еволюції насінних рослин
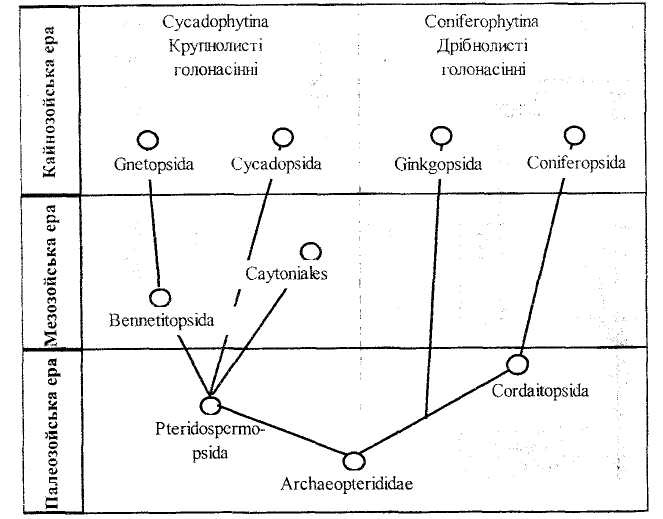
Рис. 2. Напрямки еволюції голонасінних
Табл.1. Системи голонасінних рослин
Класифікація за А.Енглером |
Класифікація за А.Тахтаджяном |
1 |
2 |
Царство Eucariota - ядерні організми Підцарство Phytobionta - рослини Відділ Telomophyta - вищі таломові рослини Підвідділ Cycadophytina - голонасінні крупно-листі |
Надцарство ядерні організми Eucariota Царство Рослини Plantae Підцарство Emdryobionta - вищі рослини Відділ Pynophyta - голонасінні |
Клас 1. Pteridospermopsida - насінні папороті Порядки: 1.1. Liginopteridales - лігіноптерисові; 1.2. Caytoniales - кейтонієві; 1.3. Glossopteridales — глосоптерисові. |
1. Клас Lyginopteridopsida насінні папороті або лігіноптеридопсиди. 1.1. Lyginopneridales - лігіноптерисові; 1.2. Caytoniales - кейтонієві; 1.3. Medullosales - медулозові; 1.4. Glossopteridales - глосоптерисові |
Клас 2. Cycadopsida - саговникові (цикадові) 2. Cycadales — саговникові |
2. Клас Cycadopsida — цикадові або саговникові 2.1. Cycadales - саговникові |
3. Bennettitopsida беннетитові Порядок 3.1. Bennettitales |
3. Клас Bennettitopsida - беннетитові, беннетитопсщи. 3.1. Bennettitales беннетитові |
Клас 4. Gnetopsida гнетові 4.1. Ephedrales - ефедрові 4 2. Welwitschiales - вельвічееві 4.3. Gnetales - гнетові |
4.Клас Gnetopsida - гнетові, гнетопсіди 4.1. Ephedrales - ефедрові 4.2. Welwitschiales - вельвічієві 4.3. Gnetales гнетові |
Підвідділ Coniferophytina - голонасінні дрібно- листі Клас 5. Ginkgopsida - гінкгові 5.1. Ginkgoales - гінкгові |
5. Клас Ginkgopsida - гінкгові, гінкгопсіди 5.1. Ginkgoales |
Клас 6.Cordaitopsida - кордаїтові Клас 7. Coniferopsida - хвойні 7.1. Taxales - тисові. Родина Тахасеае 7.2. Coniferales (Pinales) - соснові. Родини Cephalotaxaceae, Podocarpaceae, Araucariaceae, Pinaceae, Taxodiaceae, Cupressaceae |
6. Клас Pinopsida - соснові, пінопсіди Підклас Cordaitidae Порядок 6.1. Cordaitales Підклас Pinidae - хвойні або пініди 6.2. Voltziales - вольцієві 6.3. Podozamitales - подозамїтові 6.4. Araucariales - араукарієві. Родина Araucariaceae 6.5. Pinales - соснові. Родина Pinaceae 6.6. Cupressales - кипарисові. Родини Cupressaceae, Taxodiaceae 6.7. Podocarpales - подокарпові. Родина Podocarpaceae 6.8. Taxales - тисові. Родини Тахасеае, Cephalotaxaceae |
Друга лінія еволюції (рис.2), дрібнолистих голонасінних Coniferophytina від самого початку мала мало спільних з папоротеподібними ознак. Сильний приріст стебла в товщину, при слабо розвинутій серцевині (пікноксильна будова стебла), дрібні і багаточисельні листки говорять про дуже раннє походження голонасінних від різноспорових прогімноспермів з числа папоротеподібних - Archaeop-terididae.
Надалі ми будемо дотримуватися класифікації голонасінних, згідно А.Тахтаджяна, як це подано у " Жизни растений" (т. 4, 1978)
Клас Lyginopteridopsida - лігіноптеридопсіди чи насінні папороті. Появилися в девоні (360 млн. років тому - рис. 1) і вимерли в ранній крейді (140 млн. років тому). Розквіт припадає на кам'яновугільний період.
Палеоботаніки у відкладах цих геологічних періодів часто зустрічають відбитки рослин, що мали перисті листки, як у папоротей. Проте на їх жилках чи на вехівці були насінини. Через це згадані рослини в 1904 р. англійські палеоботаніки Ф.Олівер і Д.Скотт назвали насінними папоротями.
То була велика група рослин, з яких описано кілька сот видів, зведених у 4 порядки та 10 родин. Ці вимерлі рослини свого часу були поширені по всій земній кулі: котрі види більше, а котрі менше. їх місцезростаннями були заболочені приморські ліси і підвищені місця навколо них.
Ознаки лігіноптеридопсідів:
• листки папоротевого виду, але з насінинами;
• стебло теж нагадувало папороті. Були дерева, ліани і трави. Провідна система -протостела, а також сифоностела. Галуження — дихотомічне і моноподіальне. Лис-торозміщення - почергове та кільчасте (на верхівці стебла). Поверхня стебла вкрита волосками, або ще й залозками, більш-менш гладка чи вкрита основами опалих листків. Діаметр стебла - від 2-4 см до 20 см;
• насінні зачатки з пилковою камерою ( тобто порожнинами між інтегументом та нуцелусом), часто з плюскоподібним покривом (купулою), який виник із сегментів листка, що оточував зачаток;
• стробіли відсутні. Насінні зачатки розміщені поодиноко на звичайних типових листках;
• зародки у всіх досі знайдених насінинах відсутні. Є думка, що зародок розвивався після опадання насінного зачатка. Тобто ці рослини були яйцекладними. Чоловіче спороношення було як у папоротей. Мікроспори переносилися до насінного зачатка ще до повного розвитку чоловічого гаметофіту, який закінчував свій розвиток в поживній рідині пилкової камери насінного зачатка;
• продихи у насінних папоротей простогрубі, тобто гаплохейльні. Походження насінних папоротей виводять від прогімноспермних рослин, що мали вторинний ріст стебла та кореня, і були різноспоровими.
Представники класу: Медульоза Hoe (Mectallosa noei), каліматотека Хенін-гхауса (Calymmatotheca hoeninghausii) і інші.
Залишки насінних папоротей відіграли велику роль у закладанні кам'яного вугілля. Поділяється цей клас на 4 порядки і 10 родин.
Клас Cycadopsida —Цикадопсіди або Саговникові.
Це невелика група тропічних та субтропічних рослин, котрих раніше відносили до пальм, є рештками колись великої групи рослин.
Родова назва Cycas від грецького слова - пальма. Туди ж помістив їх К. Лі-ней. Життєвий цикл "від спори до спори" вивчив німецький ботанік В.Гофмейстер (1851). Всього відомо 120-130 родів, об'єднаних у 10 родів. Саговники виникли у пізньому карбоні (360 млн. років тому), розквіту набули в Юрський період (213-144 млн. років тому) і дожили до наших днів.
За сукупністю ознак вони належать до найдавніших груп голонасінних:
• листки широкі і перисті, більш - менш папоротеподібні;
• стебла мають товсту серцевину, відносно слаборозвинену і пухку вторинну ксилему та товсту кору. Тобто стебло має маноксильний тип анатомічної будови;
• спорофіли (спороносні листки) відрізняються від типових. Вони є зібраними в одностатеві стробіли, які формуються на кінчиках стебла серед листків крони.
• насінні зачатки з пилковою камерою;
• чоловічі гамети - сперматозоїди з багатьма джгутиками. Архегонії в жіночому гаметофіті дуже розвинуті.
Живуть саговники у всіх частинах світу, крім Європи та Антарктиди. Проте можна виділити три великі області їх родового ендемізму: Центральна Америка, Західна Африка з Мадагаскаром, район від Південно-Східної Азії до Австралії. Зосереджені цикадопсиди на приокеанічних територіях. Проте насінини у морській воді швидко втрачають здатність проростати. Це є основною причиною ендемізму.
Ростуть поодиноко, розсіяно, рідко невеликими групами. Частіше зустрічаються саговники в низькорослих вічнозелених склерофільних лісах і чагарникових заростях, хоч деякі види є і в тропічних дощових лісах. У загальних рисах зустрічаються вони там, де слабка міжвидова конкуренція: круті схили, скелі,
обриви.
Життєві форми (точніше форми росту) саговникових різні: розеточні дерева від 1 до 7 (20) м у висоту; кущі, а також, "безстебельні" види із вкороченим підземним стеблом; із стеблокоренем. Два види є епіфітами.
На верхівці стовбура саговникових крона із перистих листків, чим вони подібні до пальм. Вони єдині представники голонасінних з перистими листками. Листки живуть від 3 до 10 років, їх довжина від 5-6 см до 5-6 м, кількість сегментів від 4-5 до 180-200.
Є два типи стебел саговникових: підземне чи надземне бульбовидне та надземне колоновидне. Ростуть дуже повільно. Стробіли здебільшого утворюються на верхівці стебла. В подальшому верхівку формує брунька, що закладається нижче стробіла в пазусі листка.
В інших саговників стробіли закладаються під точкою росту в пазухах наймолодших листків. Звичайно, стебло не галузиться, галуження викликається механічними пошкодженнями. Відомості про максимальний вік дуже протирічні: від 100 років до 2000 років (рахують кількість крон листків на стеблі). У багатьох видів камбій функціонує протягом дуже короткого часу (роди саговник, макрозамія, бовенія), а потім закладається в корі і т.д. В інших (роди замія, цератозамія, діоона, мікроцікас) він залишається активним протягом багатьох років.
Трахеїди саговникових можуть бути спіральними, драбинчастими і пористими, причому в їх облямованих порах немає торусу. Довжина трахеїд надзвичайно велика до 7-10 мм.
Потужна кора і велика серцевина накопичують в клітинах крохмаль. Зовнішню жорсткість стеблу надають потужна кора з шарами відмерлих клітин фелеми, просочених суберином, та основи листків, що утворюють панцир.
Коренева система - стрижнева. Крім головного кореня є додаткові і також контрактильні корені. Головний корінь масивний, як стебло.
Усі саговники - строго дводомні рослини. Стробіли - дуже великі. Кількість споролистків - до 400 і навіть 600. На одній лусці мікростробіла буває до 1000 мікроспорангіїв. Це, разом з дрібним пилком, є пристосування до анемофілії. Насінних зачатків на одному споролистку мегастробіла є два. Довжина мегастро-біла максимально 1 м - у енцефаляртоса кафрського (Encephalartos caffra). Мікро-стробіли менші в кілька разів, проте є види з довгими до метра і більше мікро-стробілами (енцефаляртос вуда Encephalartos woodyi).
Вага таких великих стробілів деколи доходить до 45-50 кг (енцефаляртос кафрський), за кольором стробіли - від зеленуватих до яскравих жовтих і оранжевих. Насінні зачатки за розмірами - від 5-7 мм до 56 см.
Проростання мегаспори у саговникових дуже специфічне. Спочатку багатократно проходить каріокінез. Утворюється багато ядер ( інколи до тисячі, як у роду діоон). А тоді вже ділиться цитоплазма, тобто здійснюється цитокінез і відособлюються клітини жіночого гаметофіту. Із гаплоїдних клітин жіночого гаметофіту утворюються архегонії з яйцеклітинами, а решта клітин поступово заповнюються запасними речовинами і перетворюються в первинний ендосперм. У клітинах ендосперму саговників накопичується до 65-70 % крохмалю від загальної ваги клітини, інколи до 23 % масел, а також білки.
У клітинах жіночого гаметофіту саговникових ще зберігаються лейкопласти, вони здатні перетворитися в хлоропласти (якщо їх вийняти із мегаспорангію.)
Яйцеклітина в архегонію теж дуже велика - до 6 мм, а її ядро до 0,5 мм.
Між мікропіле і верхньою частиною гаметофіту утворюється велика порожнина. Сюди потрапляють перенесені вітром пилкові зерна. Пилок, тобто чоловічий гаметофіт, що складається з трьох клітин (вегетативної, генеративної та клітини-гаусторії) переноситься вітром. їх вловлює крапелька запилювальної рідини, що виступає з мікропіле. Пізніше вона втягується разом з пилковими зернами в пилкову камеру. Насінні зачатки виростають до розмірів насінини ще до запилення.
Від запилення до запліднення проходить півроку.
Генеративна клітина ділиться на два сперматозоїди в пилковій камері, тобто мікрогаметофіт дозріває на насінному зачатку. Сперматозоїди у саговників мають довжину до 325 мкм і є найбільшими з відомих науці.
Насінини - кулясті чи овальні мають довжину до 3-4 см і діаметр 2-3 см (найбільші з них до 8 см в довжину). Покриття насінини: епідерма, м'який шар саркотести і під ним твердий шар - склеротеста. їстівна саркотеста забезпечує зоохорію. Формування потужних насінних покривів і ендосперму ще до запилення і до запліднення - це одна архаїчна ознака. Друга ознака примітивності насінин саговникових полягає в тому, що зародок у зрілій насінині є ще недорозвинутим. Дорозвивається зародок ще протягом довгого часу після опадання насінини. Зародок збільшується в довжину в кілька разів. Ось чому разом з гінкго саговникових називають "яйцекладними" рослинами. Від розсівання насінини до проростання проходить цілий рік. Проростання насінин підземне.
На південному березі Криму розводять саговник поникаючий (Cycas revoluta) - деревце до 2-3 м у висоту. Практичне значення саговникових незначне, їх використовують аборигени в їжу. Назви "сагові пальми", "саговники", "кафрське хлібне дерево", або просто "хлібне дерево" - відображають їх значення. Мають значення цикадопсіди як декоративні і лікарські рослини. Але найбільшим є їх наукове значення, як реліктів далекого минулого нашої планети.
Клас Bennetitopsida - беннетитові або беннетитопсіди.
Група вимерлих голонасінних рослин мезозойської ери. Появилися з пермського періоду (250млн. років тому), розквіт групи прийшовся на юрський і початок крейдяного періоду. Вимерли до кінця крейдяного періоду (70 млн. років тому). Зовні були дуже подібні на сучасні і вимерлі саговники за багатьма морфологічними ознаками: будовою стебла, листків. Різниця полягає в будові епідерми і, особливо, генеративних органів.
Ознаки беннетитових:
• листки - перисті, рідше цілісні;
• продихи складногрубі - синдетохейльні;
• стебла - маноксильні;
• спорофіли зібрані у двостатеві, рідше одностатеві стробіли;
• насінні зачатки з пилковою камерою. Інтегумент витягнутий в довгу мікропілярну трубку;
• насінини за розмірами, як рисові зернятка. На відміну від насінних папоротей і саговників зародок у беннетитових був уже цілком сформований до часу опадання насіння. Крім того, у нього розвивалися 2 крупні сім'ядолі.
Запасаючих тканин нема, оскільки запасання відбувалося в сім'ядолях. Такі стробіли і таке насіння не властиві всім іншим голонасінним. А їх насінина за будовою випереджує навіть примітивні квіткові.
Представники: вільямсонія гігантська (Williamsonia gigas) — рослина з ор-тотропним стеблом до 2 м в висоту із юрської флори Йоркширу в Англії; цикадеоідея етрусків (Cycadeoidea etrusca) із Італії.
Клас Gnetopsida - Гнетові або Гнетопсіди.
Сучасні рослини. У цей клас об'єднані три роди, які, на перший погляд дуже несхожі між собою. Із ознак класу:
• листки-цілісні;
• продихи простогрубі, тобто гаплохейльні - у роду Ephedra і складногрубі - родів
Gnetum і Welwitschia;
• стебло вельвічії маноксильне, а у двох інших родів - пікноксильие. Про вельвічію можна сказати, що вона "ні на що не подібна". Гнетум скоріше нагадує тропічні дводольні деревні рослини, а ефедра подібна на австралійські дводольні рослини із роду казуаріна (Casuarina).
Незважаючи на багато розбіжностей, у цих трьох родів є спільні ознаки:
• абсолютно незвичайне для сучасних голонасінних дихазіальнє галуження стробілів;
• наявність подібного до оцвітини вкриття навколо стробілів, що також абсолютно незвичне для сучасних голонасінних;
• ознаки минулої двостатевості стробілів, особливо добре виражені у вельвічії;
• довгі мікропілярні трубки, утворені витягнутим інтегументом.
• наявність судин у вторинній ксилемі, що різко відрізняє їх від решти голонасінних, як сучасних, так і вимерлих. Крім того ще необхідно згадати загальні риси в будові оболонки пилкових зерен, двосім'ядольні зародки, супротивні листки, відсутність смоляних ходів.
Водночас вельвічія, гнетум і ефедра такі не схожі, що кожний із згаданих родів утворює цілком самостійні родини, порядки і навіть підкласи.
Рід ефедра містить більше 40 видів, розповсюджених в Європі. Азії і в Америці. Це ксерофільні рослини степів, пустель, напівпустель, піщаних і скелястих місцезростань. Здебільшого вони є низькими галузистими кущами до 6-8 м у висоту. Провідна система - ендархна еустела. Вся первинна ксилема і частина вторинної складається із трахеїд з округлими порами, що мають торус. Але у вторинній ксилемі разом з трахеїдами є справжні судини. Одначе ці судини утворилися не так як у покритонасінних. Вони виникли в результаті розширення облямованих пор на косих кінцях трахеїд і утворення там перфорацій.
Зелені та ребристі молоді стебла виконують функцію фотосинтезу. Листки лусковидні і рано опадають, розміщені супротивно або кільцями по 3 в кожному кільці.
Стробіли - одностатеві та дводомні. Зустрічаються інколи аномальні двостатеві стробіли. Зібрання мікростробілів сидять супротивно або по 3 в пазухах звичайних листків. Кожне таке зібрання складається із короткої осі, що несе від 2 до 8 супротивних пар лускоподібних покривних листків. Нижня пара стерильні, а в пазухах решти лусок сидять по одному мікростробіли. Мікростробіл має покрив з двох тонких супротивних лускоподібних листків. Всередині нього є "антерофор", що складається з колонки, на вершині якої від 2 до 8 мікросинангіїв. Ан-терофор ефедри виник в результаті зростання 2 мікроспорофілів.
Зібрання мегастробілів також сидять по 2, 3 або 4 в пазухах листків у вузлах зелених гілочок. Кожне з них складається з однієї осі, кількох пар лускоподібних листків і одного, рідше двох чи трьох, дуже редукованих мегастробілів. Кожний мегастробіл має один насінний зачаток, оточений особливим товстим і м'ясистим покривом ("оцвітиною"), який помилково називають зовнішнім інтегументом.
Інтегумент насінного зачатка (часто його звуть внутрішнім інтегументом), витягується в довгу мікропілярну трубку.
При проростанні мікроспори спочатку відбувається поділ ядер (каріокінез), але не відразу здійснюється поділ цитоплазми (цитокінез). Утворюються 2 проталі-альні ядра, генеративна клітина і клітина-трубка. Ядро генеративної клітини дає ядра сперматогенної клітини і клітини-ніжки. Із сперматогенної клітини утворяться два спермії. Таким чином, чоловічий гаметофіт редукований до шести клітин.
Із мегаспори шляхом багатократних поділів ядер і наступного формування клітинних оболонок утворюється масивний жіночий гаметофіт. В ньому закладаються 2 архегонії, кожен із 32 і більше клітин, кожен з довгою шийкою. Перенесені вітром (а навіть, можливо, ще і комахами) пилкові зерна потрапляють в крапельку рідини на мікропіле. Пізніше по мірі висихання цієї рідини, пилок втягується всередину і доходить до вершини мегаспорангію та потрапляє прямо на жіночий гаметофіт. Утворюється коротка пилкова трубка (як у хвойних і покритонасінних) і по ній спермії досягають архегоніїв. Запліднюються лише яйцеклітини.
При дозріванні насінин верхні 4 лускоподібні покривні листки мегастробілів стають соковитими і набувають жовтого чи оранжевого забарвлення. Це пристосування до зоохорії вживається також людьми під назвою "степова малина". Ефедри с лікарськими рослинами завдяки вмісту алкалоїду ефедрину.
Рід вельвічія представлений одним видом - вельвічією дивною (Welwitschia mirabilis) із пустелі Наміб. Вона - ксерофіт. Ні на що не є подібною. Головний корінь - до 3 м в довжину, стовбур маноксильний (довжина - 0.5 м, діаметр - 1.2 м) і є підсім'ядольним коліном (гіпокотілем). По краю стебла знаходяться 2 супротивних шкірястих листки, що ростуть протягом всього життя рослини - за різними даними до 2 тис. років, а може і більше. Ріст листків забезпечує інтерка-лярна меристема, довжина листків - 2-3 м, річний приріст - 8-15 см.
За анатомією стебла та морфологічною будовою всієї рослини вельвічія є дорослим проростком.
Разом з трахеїдами вельвічія має судини з простою, але неповною перфорацією на поперечних стінках і облямованими порами на бокових. Продихи на листках синдетохейльні, як у беннетитових.
Листками вона вловлює вологу з повітря. Вельвічія - дводомна рослина. Мікро- і мегастробіли утворюють складні дихазіальво галузисті зібрання над основами листків, ніби в їх пазухах. Зібрання стробілів нагадують шишки. Пилкові зерна в трьохядерному стані (проталіальна клітина, трубка, генеративна клітина) переносяться вітром. Пилок в такому трьохядерному стані з цукристою рідиною досягає нуцелуса. Генеративне ядро ділиться на 2 спермії. Цитоплазма мікрогамєтофіту не ділиться клітинними перегородками, діляться лише ядра. Цим мікрогаметофіт дуже наближається до покритонасінних.
У мегагамєтофіті теж каріокінез не супроводжується цитокінезом. Утворюються багато (до 1024) вільних ядер.
Архегонії не утворюються. При насінинах вкриття стробілів стають крилатими (анемохорія).
Рід гнетум налічує близько ЗО видів рослин із вологих тропіків, здебільшого ліан. Для них характерні великі шкірясті листки, пластинки цілісні, жилкування перистосітчасте. Подібні листки до покритонасінних дводольних.
Зібрання стробілів нагадують колоски, вони можуть галузитися. Пилок трьохклітинний (проталіальна клітина, генеративна і клітина-трубка). В пилковій трубці насінного зачатка генеративна клітина ділиться на два спермії. В гаметофіті, як у вельвічії, спочатку ядра вільні без перегородок (їх 256, рідше 512).
Після запліднення яйцеклітинами верхня частина жіночого гаметофіту набуває багатоклітинної будови.
Зародок має довгий гіпокотіль і дві сім'ядолі. Яскраво-рожеве насіння розноситься птахами.
У деяких тропічних країнах частково використовуються вироби із гнету-мів: із кори використовують міцні волокна для мотузок, молоді листки вживають в їжу, з насіння отримують олію.
Клас Гінкгових або гінкгопсіди Ginkgopsida.
Єдиний живий його представник гінкго дволопатеве (Ginkgo biloba) було відкрите для науки в 1690 р. Назва перекладається з японської як "срібний абрикос".
Розквіт цього класу припадає на кінець юрського періоду (80 млн. років
тому). Походить від насінних папоротей. Ознаки класу такі:
листки з дихотомічним жилкуванням, цілісні або з виїмкою, можуть бути дихотомічно почленованими;
продихи простогрубі - гаплохейльні;
стебла пікноксильні;
стробіли одностатеві. Рослини дводомні;
насінні зачатки з пилковою камерою;
сперматозоїди з багатьма джгутиками.
У деревині гінкго немає смоляних ходів. Листки на зиму опадають. Рослина дводомна. Мікро- і мегастробіли утворюються на вкорочених пагонах - брахі-бластах. Пилкові зерна складаються із 4 клітин: першої і другої проталіальних клітин, генеративної і клітини-гаусторії. В час запилення жіночий гаметофіт складається із гаплоїдних клітин, яких с більше 8000. При їх утворенні теж спостерігається розрив у часі між каріокінезом і цитокінезом: спочатку утворюються ядра (всі 8 тисяч), а тільки потім ділиться цитоплазма. Тканина нуцелуса адсорбується розрослим гаметофітом. До кінця червня на вершині масивного гаметофіту формуються 2 архегонії. В проміжку між ними до середини серпня появляється стовпчикоподібний виступ. Над цією ділянкою руйнується ділянка мегаспори і архегонії разом із стовпчикоподібним виступом з'являється на дні пилкової камери.
Від початку формування жіночого гаметофіту, тобто ще задовго до запліднення, в його клітинах, в т.ч. і в яйцеклітинах, накопичуються поживні речовини - крохмаль, ліпіди, білки тощо (подібно як у саговникових). Також ще задовго до запліднення (теж подібно до саговникових) у гінкго відбувається диференціація інтегументу на три шари. Внутрішній шар, ендотеста, дуже тонкий, нагадує пергаментний папір. Середній шар, склеротеста, твердий, здерев'янілий товщиною до 0,5 мм. Зовнішній шар, саркотеста, м'ясистий, товщиною 5-6 мм, покривається кутинізованою епідермою з продихами. Восени саркотеста набуває гарного жовтого кольору та неприємного запаху згірклого масла. Тобто в насінному зачатку гінкго ще до запліднення вже повністю формуються ендосперм і насінні вкриття.
Пилкові зерна від весни (часу запліднення) до осені плавають в рідині пилкової камери. Генеративна їх клітина ділиться на сперматогенну клітину і клітину-ніжку. Сперматогенна дає 2 сперматозоїди, котрі "випливають" через отвір гаусторії.
Запліднення відбувається на початку вересня, тобто через кілька місяців після запилення. Розвиток зародку, а часто і саме запліднення, у гінкго відбуваються після опадання насінних зачатків з дерева. Ця архаїчна особливість наближає гінкго до інших "яйцекладних" рослин - вимерлих насінних папоротей і кордаїтових.
Ще одна архаїчна ознака - насінина гінкго не має періоду спокою. Проростає воно десь через три місяці після запліднення, коли зародок досягне найбільшого розвитку. Проростання підземне.
У мезозойську еру рід гінкго був представлений кількома десятками видів, мав космополітний ареал, який охоплював також і територію України.
Клас Pinopsida хвойні або пінопсіди.
Це найбільш баготочисельний сучасний клас. Починається його історія із нижнього карбону (близько 360 млн. років тому ).
Із загальних особливостей класу необхідно згадати такі:
• листки цілісні з однією або двома жилками;
• продихи простогубі - гаплохейльні;
• стебла пікноксильні;
• стробічи одностатеві, утворені редукованими спорофілами.
• насінні зачатки з пилковою камерою - у підкласу кордаїтів та без пилкової камери -у підкласу соснових (хвойних);
• чоловічі гамети - спермії позбавлені джгутиків.
Поділяється клас на два підкласи: вимерлий підклас кордаїтових Cordaitidae та сучасний підклас хвойних pinidae.
Підклас Cordaitidae - представники жили від початку кам'яновугільного до кінця пермського періодів (286-248 млн. років тому). На територіях Європи та Америки в ранньому карбоні були процвітаючими. Але в різних частинах світу не водночас. Так на древньому материку Ангариді (тепер Кузбас) їх розквіт був в пермському періоді.
Кордаїти були стрункими деревами з моноподіально галузистими стеблами і високо розміщеною кроною (висота дерев 20-30 м.). Можливо то були предки сучасних хвойних, або вони мали спільне походження. V центрі стовбура була широка серцевина з поперечними розривами (подібно як у Juglandaceae).
Основну масу стовбура складала вторинна ксилема із пористих трахеїд. Листки мали довжину від кількох сантиметрів до 1 м і ширину від 1 см до 15 см. Пластинки були різної форми.
Між листками були репродуктивні пагони - стробіли, довжиною до 30 см. кожний. Але в насінинах, що були знайдені в стробілах, досі не виявлено зародків. Є гіпотеза, що зародки дуже швидко розкладалися грибами ооміцетами.
А. Енглер виділяє кордаїтів в окремий клас і разом з гінкговими і хвойними відносить до дрібнолистих голонасінних (див. табл.1.).
Підклас кордаїтів представляють роди руфлорія (Rufloria), кордайантус (Cordaianthus), кладостробус (Cladostrobus).
Підклас Pinidae - хвойні належать до найбільш знаних і найбільш важливих рослин. Вони найбільш чисельні і найбільш розповсюджені рослини серед сучасних голонасінних. Тепер їх налічується близько 560 видів, об'єднаних в 55 родів, 7 родин і 5 порядків. Величезні території Північної півкулі і частково Південної зайняті фітоценозами, де едифікаторами виступають хвойні, дуже часто один лише вид. Більшість ендемічних родів хвойних і всі древні реліктові роди зосереджені вздовж басейну Тихого океану. Причина в тому, що клімат в області Тихого океану зазнав найменших змін після мезозойської ери. По напрямку до Тихого океану збільшується і кількість видів. Наприклад, найбільше представників родів Pinus, Abies, Picea, Larix зосереджено біля нього, зокрема в Китаї.
Виникли хвойні в карбоні (370 млн. років тому), в тріасовому періоді (240 млн. років) стали відігравати відчутну роль в рослинному покриві північної півкулі, а в Юрському періоді (213-150 млн. років тому) досягай найбільшої різноманітності форм. У південній півкулі зосередилися представники родин араукарієвих (Araucariaceae) і подокарпових (Podocarpaceae), а в північній - родини соснових (Ріпасеае), таксодієвих (Taxodiaceae), тисових (Тахасеае), кипарисових (Сирressaceae). Щодо своєї древності, хвойні переважають всі нині живучі групи голонасінних. Дивно, але древній вік мають і окремі роди: араукарія (Araucaria) - 200 млн. років, кедр (Cedrus) - 250 млн. років, сосна - 135 млн. років. Серед хвойних рекордсмени по віку і за розмірами стовбура такі: висота 1 12 метрів у секвої вічнозеленої (Sequoia sempervirens), 117м у псевдотсуги Мензіса (Psendotsuga menziesii); діаметр 12 м у секвоядендрона гігантського (Sequiadendron sempervirens), 11 м - у секвої вічнозеленої, 16 м у такодіума мексіканського (Taxodium mucro-natum); 3000 років - вік окремих екземплярів секвойядендрона і 4900 років сосни довгоживучої (Pmus longaeva).
Серед цього підкласу зустрічаємо дерева і чагарники найрізноманітніших форм та розмірів. Найменші розміри у сланкого виду дакрідіума рихлолистого (Dacrydium laxifolium) із Нової Зеландії, прозваного "сосною-пігмесм".
Галуження стебла хвойних моноподіальне. Гілки зібрані часто у стиснуті спіралі, що нагадують мутовки (у сосен, ялин) і по них можна визначати вік дерев. При пошкодженні верхівки одна з бокових гілок починає рости вверх і формує верхівку. У старих дерев буває відразу по декілька вершин, через що крона стає зонтиковидною. У багатьох представників підкласу є не один, а два типи пагонів: ауксібласти і брахібласти. Бруньки з лусками або без них.
Анатомічна будова стебла хвойних досить одноманітна: слабо розвинута серцевина, масивний циліндр деревини, відносно тонка кора. Провідна система - ендархна евстела. Ксилема по об'єму на 90-95 % складається із трахеїд, довжина котрих від 0,5 до 11 мм. На радіальних і поперечних стінках трахеїд розміщені облямовані пори з торусом. Паренхіми дуже мало, або вона відсутня. Розміщена паренхіма навколо смоляних ходів, зрідка поміж трахеїдами. Серцевинні промені складаються або тільки з паренхімних клітин, або мають ще по 1 -2 ряди променевих трахеїд. Характер полів перехресту клітин променів з поздовжніми трахеїдами є важливою діагностичною ознакою систематичної приналежності деревини.
У деревині і в корі багатьох хвойних є багато смоляних ходів (смоляних каналів).
У більшості хвойних у стовбурі чітко виражені річні кільця приросту деревини. Первинний корінь у більшості хвойних зберігається протягом всього життя і розвивається у потужний стержень, від якого відходять бокові корені. На поглинаючих корінцях часто розвинута мікориза.
Листки хвойних дрібні і в сучасних видів завжди цілісні. У багатьох хвойних листки двох типів: зелені фотосинтезуючі та безхлорофільні коричневі лускоподібні. Найбільші листки у подокарпа найбільшого (Podocarpus maximus) довжина до 35 см і ширина листків до 9 см, та в сосни болотної довжина до 45 см. Вузькі листки мають одну нерозгалужену жилку, або ця одна жилка роздвоєна (рід Pinus підрід Pinus або Diploxylon). Широкі листки мають кілька паралельних жилок, при основі дихотомічно розділених. Листки багаторічні (крім родів Модрина, псевдомодрина, метасеквойя, таксодіум), шкірясті і жорсткі. Твердості їм надає гіподерма, що складається з 1-3 рядів склеренхіми і утворює таким чином твердий зовнішній скелет.
Стробіли хвойних завжди одностатеві. В результаті редукції із первинних викопних сережкоподібних зібрань стробілів збереглися у сучасних хвойних лише поодинокі стробіли, що відповідають верхівковому мікростробілу в "сережках" підкласу кордаїтових.
Мікростробіл (що часто зветься "чоловічим колоском") складається із вкороченої осі, на якій розміщені сильно редуковані мікроспоролистики. У родини кипарисових вони розміщені завжди супротивно або мутовчасто, у всіх інших хвойних спірально. Є два типи мікроспорофілів: дорсовентральний (як у сосни) та радіально-симетричний (як у тису).
Мегастробіли, як і мікростробіли, є продуктом спеціалізації сережкоподіб-них зібрань стробілів предкових форм. Число мегастробілів зменшилося до одного. Зменшення мегастробілів у тису призвело до утворення поодиноких однонасінних мегастробілів.
У більшості хвойних еволюція мегастробілів йшла шляхом скорочення довжини міжвузлів головної осі і утворення компактного зібрання мегастробілів, яке ми звемо жіночою шишкою, або просто шишкою.
Кожна шишка (наприклад сосни, ялини та ін.) складається із центральної осі, на якій сидять покривні луски (видозмінені листки), в пазухах кожної з них знаходиться так звана "насінна луска". їх звуть "насінними", бо кожна на своїй верхній стороні несе насінні зачатки.
Насінна луска - це не листовий орган, не спорофіл, тим більше не плодолистик. Насінна луска — це видозмінений і спеціалізований брахібласт (вкорочений пагін), розміщений в пазусі покривної луски.
Із розміщенням насінних зачатків на верхній стороні насінної луски появилася тенденція до її зростання з покривною лускою. В напрямку від родини соснових через ряд проміжних форм до кипарисових проходить зростання насінної луски з покривною.
Надзвичайно своєрідною у хвойних є форма пилкових зерен (пилку). У більшості хвойних вони за формою нагадують біб з двома повітряними мішками. У тсуги пилкові зерна кулясті з одним повітряним мішком, у модрини і псевдотсуги кулясті без повітряних мішків.
Розвиток пилкових зерен (тобто чоловічого гаметофіту) починається ще до розкривання пиляків. В результаті мітотичних поділів мікроспори утворюються дві чи більше маленькі проталіальні клітини і одна велика антеридіальна, яка ділиться на маленьку генеративну і більшу клітину-трубку. Генеративна клітина ділиться на стерильну клітину-ніжку і сперматогенну клітину, що дає два спермії. Спермії утворюються перед заплідненням на жіночому гаметофіті, вони не мають джгутиків.
Подібно до інших голонасінних насінний зачаток хвойних складається із інтегумента, нуцелуса (мегаспорангія) та розвинутого в ньому жіночого гаметофіта.
На верхівці жіночого гаметофіа, що складається з багатьох (близько 1000) клітин, утворюються архегонії з крупними яйцеклітинами. Всі клітини жіночого гаметофіту хвойних, крім клітин архегоніїв, запасають поживні речовини і ще до запліднення формують ендосперм (первинний ендосперм). Насінини опадають, маючи цілком сформований зародок. Тобто хвойні є "живородящими" рослинами. Запліднення у хвойних починається зразу після запилення, і лише у роду сосна між цими процесами проходить 12-14 місяців.
Зародок насінини з клітин підвіски, зародкового корінця, стебельця і сім'ядолей (числом від 3 до 18).
Згідно зі загальноприйнятою системою хвойних, запропонованої голландським ботаніком А.Пулле (1937, 1950), в підкласі хвойних виділяють 7 порядків вимерлі порядки вольцієвих (Voltziales) і подозамітових (Podozamitales); сучасні 5 порядків - араукарієвих (Araucariales), подокарпових (Podocarpales), соснових (Pinales), кипарисових (Cupressales), тисових (Taxales).
1. Представники порядку вольцієвих мали багато спільного з кордаїтами. їх геологічна історія тривала від пізнього карбону до ранньої крейди (300-140 млн. років тому). Трахеїди - з 1-3 рядами пор (подібно до араукарієвих), смоляні ходи відсутні. Листки - почергові, диморфні (неоднакові на стовбурах та гілках), розміщені спірально, широкі чи голчасті. Жилкування - просте. Мікростробіли - на кінцях бокових гілочок. Кожний мікроспофіл з 2 пиляками на нижній стороні. Пилкові зерна - з одним суцільним пилковим мішком. Рід ульманія (Ullmannia) та інші.
2. Представники порядку подозамітових (Podozamitales) існували від пізнього тріасу до пізньої крейди (250-70 млн. років тому). Листки почергові, відносно крупні, з пластинками різної форми. Шишки з недерев'яніючими листоподібними покривними лусками і невеликими вільними, часто роздільними насінними лусками з 2-4 насінними зачатками. Представлений порядок однією родиною Podozamitaceae.
3. Порядок араукарієвих (Araucariales) відомий від пізньої пермі (250 млн. років тому). Характерною ознакою деревини є араукаріодна пористість: облямовані пори на стінках трахеїд скупчені, здебільшого стикуються і змикаються, внаслідок чого стають шестикутними. Смоляні ходи зовсім відсутні. Паренхіми в деревині немає. Листки - почергові, більш-менш крупні, плоскі, різної форми до шиловидних. Жилкування - дихотомічне, паралельне, а в шиловидних листках - просте. Мікростробіли - великі із багаточисельних розміщених спірально споролистиків, що мають по 5-20 пиляків. У пилковому зерні від ІЗ до 40 проталіаль-них клітин. Насінні і покривні луски шишок майже повністю зрослися, на кожній лусці по одному насінному зачатку. В порядку одна родина араукарієвих Araucariaceae, що включає два роди: агатис (Agatis) і араукарія (Araucaria).
4. Порядок соснових (Pinales) відомий з тріасового періоду (250 млн. років). Облямовані пори на стінках трахеїд однорядні, рідше двохрядні. У багатьох представників постійно є смоляні ходи. Паренхіми в деревині мато, або вона відсутня. Листки - почергові, голчасті. Мікроспорофіли - дорсовентральні, розміщені спірально, кожний з 2 пиляками. Пилкові зерна у більшості родів з 2 повітряними мішками. Чоловічий гаметофіт - з двома проталіальними клітинами. Шишки - із спірально розміщених покривних і насінних лусок. На лусці 2 насінних зачатки. Насінина - з крилом, яке утворюється із поверхневих шарів насінної луски. Зародок з 3-18 сім'ядолями. В порядку 1 родина Ріпасеае, що включає 11 родів і більше 250 видів.
5. Порядок кипарисові Cupressaceae. Його геологічна історія почалася в юрському періоді (200 млн. років тому). Поровість трахеїд - однорядна. Смоляні ходи відсутні. Паренхіми в ксшіемі є досить багато. Листки - почергові, супротивні і мутовчасті; лінійно-ланцетні, голчасті та лусковидні з однією жилкою. Мікростробіли дрібні, мікроспорофіли розміщені як і листки, несуть по 2-9 пиляків. Пилкові зерна - без повітряних мішків, без проталіальних клітин. Шишки зі зрослими покривними та насінними лусками. В основі луски від 2 до 12 насінних зачатків. Насінини крилаті або безкрилі.
У складі порядку дві родини: таксодієвих (Taxogiaceae - 10 родів, 14 видів) і кипарисових (Cupressaceae - 19 родів., 130 видів).
6. Порядок Taxales - тисових появився в кінці тріасового періоду (220 млн. років тому). Трахеїди - з однорядною поровістю і великими третинними потовщеннями стінок. Смоляні ходи відсутні. Паренхіми в деревині багато, мало або зовсім немає. Листки - почергові, лінійні або ланцетні. Мікроспорофіли з 2-8 пиляками. Пилок - без повітряних мішків, без проталіальних клітин. Мегастробіли - поодинокі з одним насінним зачатком. В основі насінного зачатку є подібний до комірця покрив - аріллюс, що відповідає насінній лусці. Зародок з двома сім'ядолями. В цьому порядку є дві сучасних родини: тисових (Тахасеае - 5 родів) і головчатотисових (Cephalotaxaceae - 1 рід, 6 видів).
7. Порядок Podocarpaies подокарпових виник в пермському періоді (280 млн. років тому). Трахеїди - з одним рядом пор. Смоляні ходи відсутні. Листки почергові або супротивні, від широколанцетних до лусковидних. У деяких видів роду подокарп (Podocarpus) - великі і широкі листки. Мікростробіли дрібні, на мікроспоролистиках - по 2 пиляки. Пилкові зерна мають 8 проталіальних клітин. Типової шишки немає. Мегастробіл має один насінний зачаток, оточений сильно редукованою насінною лускою - епімантієм. Зародок з двома сім'ядолями. Порядок представлений однією сучасною родиною подокарпових (Podocarpaceae - 140 видів), що зростає у вологих районах південної півкулі.