


М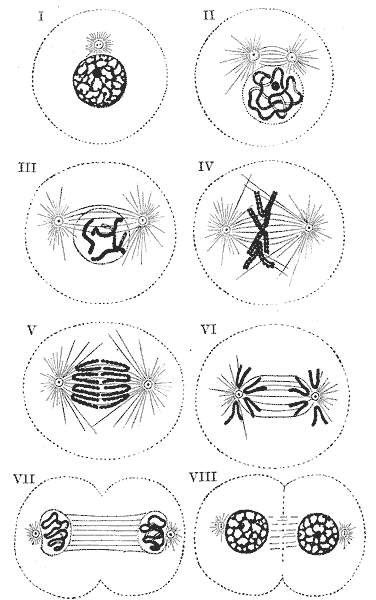 ітоз
(рідше: каріокінез або непрямий поділ)
— поділ ядра еукаріотичної клітини зі
збереженням числа хромосом. На відміну
від мейозу, мітотичний поділ протікає
без ускладнень в клітинах будь-якої
плоїдності, оскільки не включає як
необхідний етап кон'югацію гомологічних
хромосом протягом профази.
ітоз
(рідше: каріокінез або непрямий поділ)
— поділ ядра еукаріотичної клітини зі
збереженням числа хромосом. На відміну
від мейозу, мітотичний поділ протікає
без ускладнень в клітинах будь-якої
плоїдності, оскільки не включає як
необхідний етап кон'югацію гомологічних
хромосом протягом профази.
У 1874 І. Д. Чистяков описав ряд стадій (фаз) мітозу у спорах плаунів, ще не ясно представляючи собі їхню послідовність. Детальні дослідження з морфології мітозу уперше були виконані Е. Страсбургером на рослинах (1876 — 1879) і В. Флеммінгом на тварин (1882).
Фази мітозу:
У профазі відбувається конденсація хромосом, клітинний центр ділиться і продукти його поділу розходяться до полюсів ядра, руйнується ядерна оболонка, утворюється веретено поділу.
У
I — III — профаза; IV — метафаза; V — VI — анафаза; VII — VIII — телофаза.
метафазі хромосоми розташовуються на «екваторі» (на рівній відстані від «полюсів» ядра) в одній площині, утворюючи так звану метафазну пластинку. Важливо відзначити, що вони залишаються в такому положенні протягом досить тривалого часу. Зазвичай у зв'язку з цим метафаза — найслушніший час для підрахунку хромосомних чисел.У анафазі, яка є найкоротшою фазою мітозу, хромосоми діляться (з'єднання в районі центромери руйнується) і розходяться до різних полюсів клітини.
У тілофазі відбувається руйнування веретена поділу і утворення ядерної оболонки навколо дочірніх ядер.
Варіанти мітозу
Слід зазначити, що в різних групах живих організмів мітоз протікає дещо по-різному. Описаний вище варіант мітозу називається відкритий ортомітоз (ядерна оболонка руйнується, веретено поділу пряме, оскільки продукти поділу клітинного центру розміщені на протилежних полюсах ядра). Характерний для багатоклітинних тварин, багатоклітинних рослин і ряду найпростіших.
У деяких групах найпростіших продукти поділу клітинного центру в анафазі не досягають протилежних сторін ядра, внаслідок чого мікротрубочки веретена поділу розташовуються під кутом, нагадуючи букву V (такий варіант ділення отримав назву плевромітоз). У ряді випадків мітоз відбувається без руйнування ядерної оболонки (закритий мітоз). Нарешті, клітинний центр може містити центріолі (наприклад, у тварин) або не містити їх (наприклад, у квіткових рослин). Відповідно, розрізняють також центріолярний і ацентріолярний мітоз.
Варіанти класифікації мітозів
7 типів мітозу найпростіших:
Закритий внутрішньоядерний плевромітоз
Закритий внутрішньоядерний ортомітоз
Закритий евгленоідний мітоз
Закритий позадерний плевромітоз
напівзакритий плевромітоз
напівзакритий ортомітоз
Відкритий ортомітоз (еумітоз)
6 типів мітозу водоростей:
Закритий центрический
Закритий ацентріческій
Напівзакритий центрический
Напівзакритий ацентріческій
Відкритий центрический
В
 ідкритий
ацентріческій
ідкритий
ацентріческій
Підготовка до поділу
Еукаріотичні організми, що складаються з кліток, що мають ядра,
починають підготовку до розподілу на визначеному етапі клітинного циклу,
в інтерфазі.
Саме в період інтерфази в клітці відбувається процес біосинтезу білка,
подвоюються всі найважливіші структури клітки. Уздовж вихідної хромосоми
з наявних у клітці хімічних сполук синтезується її точна копія,
подвоюється молекула ДНК. Подвоєна хромосома складається з двох
половинок - хроматид. Кожна з хроматид містить одну молекулу ДНК.
Інтерфаза в клітках рослин і тварин у середньому продовжується 10 - 20
годин. Потім настає процес розподілу клітки - мітоз.
Під час мітозу клітка проходить ряд послідовних фаз, у результаті яких
кожна дочірня клітка одержує такий же набір хромосом, який був у
материнській льотці.
Поділ кліток здійснюється поетапно. На кожному етапі поділу відбуваються
визначені процеси. Вони приводять до подвоєння генетичного матеріалу
(синтезу ДНК) і його розподілу між дочірніми клітками. Період життя
клітки від одного поділу до наступного називається клітинним циклом.
Фази мітозу
Профаза
До основних подій профази відносять конденсацію хромосом усередині ядра і утворення веретена поділу в цитоплазмі клітини. Розпад ядерця в профазі є характерною, але не обов'язковою для всіх клітин особливістю.
Умовно за початок профази приймається момент виникнення мікроскопічно видимих хромосом внаслідок конденсації внутрішньоядерної хроматину. Ущільнення хромосом відбувається за рахунок багаторівневої спирализации ДНК. Дані зміни супроводжуються підвищенням активності фосфорілаз, модифікуючих гістони, безпосередньо беруть участь у компонуванні ДНК. Як наслідок, різко знижується транскрипційних активність хроматину, інактивуються ядерцеві гени, велика частина ядерцевих білків дисоціюють. Конденсуються сестринські хроматиди в ранній профазі залишаються спареними по всій своїй довжині за допомогою білків-когезінов, однак до початку прометафаза зв'язок між хроматида зберігається лише в області центромер. До пізньої профазі на кожній Центромера сестринських хроматид формуються зрілі кінетохори необхідні хромосомам для приєднання до микротрубочкам веретена поділу в прометафазі.
Поряд з процесами внутрішньоядерної конденсації хромосом в цитоплазмі починає формуватися мітотичний веретено - одна з головних структур апарату клітинного ділення, відповідальна за розподіл хромосом між дочірніми клітинами. В утворенні веретена розподілу у всіх еукаріотичних клітин беруть участь полярні тільця (центросоми), мікротрубочки і кінетохори хромосом.
З початком формування мітотичного веретена в профазі сполучені разючі зміни динамічних властивостей мікротрубочок. Час напівжиття середньої мікротрубочки зменшується приблизно в 20 разів від 5 хвилин (в інтерфазі) до 15 секунд. Однак швидкість їх росту збільшується приблизно в 2 рази в порівнянні з тими ж інтерфазних мікротрубочками. Полімеризується плюс-кінці («+»-кінці) є «динамічно нестабільними» і різко переходять від рівномірного зростання до швидкого укороченню, при якому часто деполімеризує вся микротрубочка. Примітно, що для правильного функціонування мітотичного веретена необхідний певний баланс між процесами зборки і деполімеризації мікротрубочок, так як ні стабілізовані, ні деполімеризований мікротрубочки веретена не в змозі переміщати хромосоми.
Паралельно конденсації хромосом і формування мітотичного веретена, під час профази відбувається фрагментація ендоплазматичного ретикулуму, який розпадається на дрібні вакуолі, що розходяться потім до периферії клітини. Одночасно рибосоми втрачають зв'язку з мембранами ЕПР. Цистерни апарата Гольджі також міняють свою околоядерном локалізацію, розпадаючись на окремі діктіосоми, без особливого порядку розподілені в цитоплазмі.
Прометафаза
Закінчення профази і наступ прометафаза, як правило, знаменується розпадом ядерної мембрани. Цілий ряд білків ламіни фосфорилюється, внаслідок чого ядерна оболонка фрагментируется на дрібні вакуолі, а порові комплекси зникають. Після руйнування ядерної мембрани хромосоми без особливого порядку розташовуються в області ядра. Однак незабаром всі вони приходять в рух.
У прометафазі спостерігається інтенсивне, але безладне переміщення хромосом. Спочатку окремі хромосоми стрімко дрейфують до найближчого полюсу мітотичного веретена зі швидкістю, що досягає 25 мкм / хв. Поблизу полюсів ділення підвищується ймовірність взаємодії новосинтезованих плюс-решт мікротрубочок веретена з кінетохорамі хромосом. У результаті такої взаємодії кінетохорние мікротрубочки (пов'язані з кінетохором) стабілізуються від спонтанної деполімеризації, а їх зростання частково забезпечує віддалення з'єднаної з ними хромосоми в напрямку від полюса до екваторіальній площині веретена. З іншого боку хромосому наздоганяють тяжі мікротрубочок, що йдуть від протилежного полюса мітотичного веретена. Взаємодіючи з кінетохором, вони також беруть участь в русі хромосоми. В результаті сестринські хроматиди виявляються пов'язаними з протилежними полюсами веретена. Зусилля, що розвивається мікротрубочками від різних полюсів, не тільки стабілізує взаємодія цих мікротрубочок з кінетохорамі, але також, у кінцевому рахунку, призводить кожну хромосому в площину метафазних платівки.
У клітинах ссавців прометафаза протікає, як правило, протягом 10-20 хвилин.
У нейробластів коника дана стадія займає всього 4 хвилини, а в ендоспермі Haemanthus і в фібробластах тритона - .. Близько 30 хвилин. У дріжджових клітинах неможливо чітко розмежувати стадії профази і прометафаза з причини збереження ядерної оболонки в процесі ділення. Аналогічним чином, часткове або більш пізніше руйнування ядерної мембрани ускладнює розмежування стадій профази і прометафаза в клітинах Drosophila і C. Елеганс. У подобру випадках для опису всіх ранніх подій мітотичного поділу використовується узагальнюючий термін «профаза».