

Задание. 1. Изучить представленный теоретический материал.
2. Написать конспект лекций.
3. Рассмотреть все иллюстрации, подобрать (придумать) примеры для каждой
из них.
Лекция . Факторы микроэволюции.
Количественная характеристика естественного отбора. Для количественной характеристики отбора обычно используется относительная приспособленность, называемая также адаптивной или селективной ценностью генотипа, под которой понимается способность генотипа к выживанию и размножению. Приспособленность обозначается буквой w и колеблется в пределах от 0 до 1. При w = 0 передача наследственной информации следующему поколению отсутствует из-за гибели всех особей; при w = 1 полностью реализуются потенциальные возможности к размножению. Величина, дополняющая приспособленности генотипа, называется коэффициентом интенсивности отбора и обозначается буквой S: w + S = 1, отсюда
S = 1 — w,
w = 1 — S.
Коэффициент интенсивности отбора определяет скорость уменьшения частоты того или иного генотипа. Чем больше коэффициент интенсивности отбора и чем, следовательно, меньше приспособленность каких-либо генотипов, тем выше давление отбора.
 Генетический
дрейф. Возрастание
численности вида, как правило,
сопровождается расширением его ареала.
Попав за пределы ареала,
небольшие популяции могут закрепиться
в новых местах обитания и тогда
подвергнутся действию условий, совершенно
отличающихся
от таковых в пределах ареала вида. В
таких периферических
микропопуляциях влияние случайных
факторов может выйти
на первый план и привести к резкому
изменению генофонда.
Генетический
дрейф. Возрастание
численности вида, как правило,
сопровождается расширением его ареала.
Попав за пределы ареала,
небольшие популяции могут закрепиться
в новых местах обитания и тогда
подвергнутся действию условий, совершенно
отличающихся
от таковых в пределах ареала вида. В
таких периферических
микропопуляциях влияние случайных
факторов может выйти
на первый план и привести к резкому
изменению генофонда.
Случайное ненаправленное изменение частот аллелей в малых популяциях Н. П. Дубинин и Д. Д. Ромашов (1931 —1932) назвали генетико-автоматическими процессами. Независимо от них зарубежные ученые С. Райт и Р. Фишер назвали это явление генетическим дрейфом.
Генофонд малочисленной популяции с самого начала беднее генофонда родительского вида, и подбор аллелей в нем случаен (в результате случайного подбора особей). Поскольку естественный отбор происходит эффективно при достаточно большой численности особей, дальнейшая судьба генофонда малой популяции определяется главным образом действием различного рода случайных факторов. Это было доказано экспериментально. В пробирки с кормом посадили по две самки и два самца мух дрозофил, гетерозиготных по аллелю А (из генотипа Аа). В такой искусственно созданной популяции соотношение нормального и мутантного аллелей было одинаковым. Спустя несколько поколений оказалось, что частота мутантного аллеля меняется случайным образом. В некоторых популяциях он был утрачен, в других, наоборот, все особи стали гомозиготными по мутантному аллелю, часть популяций содержала нормальный и мутантный аллели.
 Таким
образом, несмотря на снижение
жизнеспособности мутантных
особей и, следовательно, вопреки
естественному отбору в
некоторых небольших популяциях мутантный
аллель полностью вытеснил
нормальный. Это и есть результат
случайного процесса — дрейфа генов.
Таким
образом, несмотря на снижение
жизнеспособности мутантных
особей и, следовательно, вопреки
естественному отбору в
некоторых небольших популяциях мутантный
аллель полностью вытеснил
нормальный. Это и есть результат
случайного процесса — дрейфа генов.
Соотношение роли отбора и дрейфа генов при разной численности популяций можно выразить следующим образом: если численность популяции N< 1|2S, то преобладает воздействие дрейфа генов, если N>1|4S, то преобладает отбор, в интервале между этими величинами действуют совместно оба фактора.
Изоляция. Под изоляцией понимается возникновение любых барьеров, нарушающих панмиксию. Еще Ч. Дарвин придал изоляции большое значение в видообразовании. Но он рассматривал явления, связанные с изоляцией, в большом, макроэволюционном масштабе. Нас же изоляция интересует как элементарный эволюционный фактор, действующий вначале в пределах популяции, на микроэволюционном уровне.
В зависимости от природы изолирующих барьеров выделяют два основных типа изоляции: географическую и биологическую, или репродуктивную.
Географическая изоляция. Географическая изоляция связана с изменениями в ландшафте: образование преград в виде рек, горных хребтов, лесных массивов и т. д. В результате свободное скрещивание сухопутных особей затрудняется из-за водных барьеров, а особей, обитающих в воде, — из-за барьеров суши. Возвышенности изолируют равнинные популяции, а равнины — горные популяции.
Географическая изоляция может возникнуть и без заметных географических барьеров, просто в силу большого расстояния между популяциями одного вида. Часто причиной такой изоляции является деятельность человека в биосфере. Например, еще в начале нашего века ареал соболя был сплошным и занимал значительную часть Евразии, но уже в 20—30-х годах в связи с интенсивным перепромыслом этого зверька его ареал приобрел мозаичную структуру: распался на отдельные участки, разделенные значительным расстоянием, препятствовавшим свободному скрещиванию удаленных друг от друга популяций. В 40—50-е годы благодаря большой работе охотоведов по восстановлению численности соболя и расселению зверьков в местах их былого обитания ареал соболя в СССР был восстановлен.
 Степень
географической изоляции. Изоляция
может быть абсолютной.
Например, популяции песцов, обитающие
на Командорских островах и на Камчатке,
не обмениваются особями, так как разделены
незамерзающим проливом. Однако изоляция
зависит не только от пространственного
расположения географических барьеров,
но и от радиуса
индивидуальной активности —
расстояния, на которое способны
перемещаться особи в период размножения.
Если радиус индивидуальной активности
мал по
сравнению с размерами популяции и
расстоянием между соседними
популяциями, то степень,
или давление, изоляции будет
большим.
Например, популяции наземных моллюсков,
удаленные друг
от друга на 200—400 м, будут существовать
изолированно, так как их индивидуальная
подвижность ограничена. А песцы
могут мигрировать по льдам Арктики на
сотни километров, и в результате
изоляция песцов острова Врангеля от
популяций песцов Чукотки очень
невелика. Если радиус индивидуальной
активности
позволяет особям соседних популяций
преодолевать расстояние между ними
(степень изоляции незначительная), то
эти популяции очень скоро приобретут
общие свойства из-за полного перемещения
генетического материала. Так, утки
одного вида занимают
ареал, равный чуть не всей Европе, и
свободно скрещиваются между собой.
Предполагают, что все они относятся к
одной популяции
или очень немногим популяциям.
Степень
географической изоляции. Изоляция
может быть абсолютной.
Например, популяции песцов, обитающие
на Командорских островах и на Камчатке,
не обмениваются особями, так как разделены
незамерзающим проливом. Однако изоляция
зависит не только от пространственного
расположения географических барьеров,
но и от радиуса
индивидуальной активности —
расстояния, на которое способны
перемещаться особи в период размножения.
Если радиус индивидуальной активности
мал по
сравнению с размерами популяции и
расстоянием между соседними
популяциями, то степень,
или давление, изоляции будет
большим.
Например, популяции наземных моллюсков,
удаленные друг
от друга на 200—400 м, будут существовать
изолированно, так как их индивидуальная
подвижность ограничена. А песцы
могут мигрировать по льдам Арктики на
сотни километров, и в результате
изоляция песцов острова Врангеля от
популяций песцов Чукотки очень
невелика. Если радиус индивидуальной
активности
позволяет особям соседних популяций
преодолевать расстояние между ними
(степень изоляции незначительная), то
эти популяции очень скоро приобретут
общие свойства из-за полного перемещения
генетического материала. Так, утки
одного вида занимают
ареал, равный чуть не всей Европе, и
свободно скрещиваются между собой.
Предполагают, что все они относятся к
одной популяции
или очень немногим популяциям.
Биологическая, или репродуктивная, изоляция. Биологическая (репродуктивная) изоляция приводит к нарушению скрещивания или препятствует воспроизведению нормального потомства. Различают несколько форм репродуктивной изоляции.
 При
эколого-этологической
изоляции свободное
скрещивание между организмами нарушается
в результате снижения вероятности
встреч партнеров из разных популяций
в период размножения. Причиной этого
могут быть различия в образе жизни и
поведении. Примеров такого рода много.
Например, у птиц одного вида могут
возникать различия в сроках спаривания,
ритуале ухаживания, брачных песнях
и т. д. У растений одного вида многочисленны
случаи несовпадения времени цветения
и опыления. В озере Севан существовало
несколько популяций форели, отличающихся
размерами, темпами роста, сроками и
местами нереста. Однако эта форма
изоляции
не всегда надежное препятствие
к скрещиванию. Имеется
немало случаев нарушения экологических
и поведенческих изоляционных барьеров.
При
эколого-этологической
изоляции свободное
скрещивание между организмами нарушается
в результате снижения вероятности
встреч партнеров из разных популяций
в период размножения. Причиной этого
могут быть различия в образе жизни и
поведении. Примеров такого рода много.
Например, у птиц одного вида могут
возникать различия в сроках спаривания,
ритуале ухаживания, брачных песнях
и т. д. У растений одного вида многочисленны
случаи несовпадения времени цветения
и опыления. В озере Севан существовало
несколько популяций форели, отличающихся
размерами, темпами роста, сроками и
местами нереста. Однако эта форма
изоляции
не всегда надежное препятствие
к скрещиванию. Имеется
немало случаев нарушения экологических
и поведенческих изоляционных барьеров.
При морфофизиологической изоляции изменяется не вероятность встречи полов, а вероятность оплодотворения. Это может быть обусловлено различием в строении полового аппарата, нарушением процессов размножения.
Очень важная форма биологической изоляции — генетическая изоляция. К ней относятся все те случаи, когда в результате скрещивания форм с разными хромосомными наборами появляются гибриды с пониженной жизнеспособностью, плодовитостью или полной стерильностью.
Эволюционное значение изоляции. Различные формы изоляции, нарушая панмиксию и действуя длительное время, закрепляют различия в частотах встречаемости различных генотипов (тогда как свободное скрещивание ведет к сглаживанию различий). Однако, несмотря на длительность действия изоляции, ее давление на популяцию так же ненаправленно, как и давление факторов (мутационные процессы, «волны жизни») — поставщиков эволюционного материала.
Ненаправленность названных факторов эволюционного процесса — главная черта, отличающая их от естественного отбора — направленного элементарного эволюционного фактора. Факторы-поставщики эволюционного материала изменяют частоты аллелей, а изоляция усиливает их действие независимо от того, ведет это к повышению или к понижению приспособленности организмов к условиям существования.
Естественный отбор с точки зрения генетики. Сущность естественного отбора заключается не столько в выживании наиболее приспособленных, сколько в гибели неприспособленных особей или уменьшении вклада последних в генофонд следующих поколений. Сам же факт выживания, вплоть до достижения старости без оставления потомства, не будет иметь последствий для эволюции. Поэтому естественный отбор определяют как процесс, направленный к повышению (или понижению) вероятности оставления потомства одними особями по сравнению с другими.
Отбор происходит внутри популяции, между популяциями и между видами. Нас интересует прежде всего внутрипопуляционный отбор, так как именно он является элементарным эволюционным фактором. В пределах популяции отбираются (преимущественно оставляют потомство) не отдельные признаки организма, а особи, обладающие ими. Следовательно, отдельные особи — это элементарный объект отбора.
Совокупность внешних признаков особи — ее фенотип представляет собой результат взаимодействия генотипа с условиями среды. Отбор всегда идет по фенотипам. Но в фенотипе особи отражаются особенности генотипа. Вот почему в ряду поколений отбор по фенотипам сводится к отбору генотипов с нормой реакции, соответствующей условиям данной среды. Таким образом, под естественным отбором следует понимать избирательное воспроизведение разных генотипов.
Отбор и мутации. Особенно эффективно отбор идет против доминантных мутаций, поскольку они проявляются сразу не только в гомозиготном, но и в гетерозиготном состоянии. При S=l популяция за одно поколение избавляется от доминантных летальных мутаций. Например, доминантным аллелем обусловлено тяжелое заболевание у человека — ахондроплазия. Из-за нарушения роста длинных костей для таких больных характерны короткие, часто искривленные конечности и деформированный череп. Гомозиготы по этому аллелю совершенно нежизнеспособны (S = 1). У гетерозигот число детей впятеро меньше по сравнению со здоровыми людьми, т. е. w = 0,2; S = 0,8.
В качестве доминантных мутаций можно рассмотреть и некоторые хромосомные перестройки. Так, больные с синдромом Дауна, как правило, не оставляют потомства (S = 1), и популяция избавляется от этого вредного гена за одно поколение. Но почему тогда заболевания, обусловленные доминантными мутациями, не исчезают бесследно? Это объясняется непрерывным действием мутационного процесса, поддерживающего присутствие вредных аллелей в популяции. Так, частота возникновения аллеля ахондроплазии равна 1 на 20 000 гамет, т. е. частота новорожденных детей с этой болезнью в потомстве здоровых родителей составит 1:10000. Многие рецессивные мутации обладают пониженной приспособленностью и будут устраняться отбором. Если рецессивные гомозиготы обладают нулевой приспособленностью, то популяция избавится от них также за одно поколение. Но отбор против рецессивных аллелей затруднен, потому что большая их часть находится в гетерозиготном состоянии (под прикрытием нормального фенотипа) и они как бы ускользают от действия отбора. Подсчитано, что если частота «вредного» рецессивного аллеля равна 0,01, то потребуется 100 поколений только для того, чтобы снизить частоту аллеля вдвое, и 9900 поколений, чтобы снизить ее до 0,0001. Особенно трудно избавить от рецессивных мутаций большие популяции, так как в них вероятность перевода таких мутаций в гомозиготное состояние очень мала.
 Отбор
в пользу гетерозигот. Нередко
наблюдается отбор в пользу
гетерозигот, когда обе гомозиготы имеют
пониженную приспособленность
по сравнению с гетерозиготами. Хорошо
известным примером
такого отбора в популяциях человека
является серповидноклеточная анемия
— болезнь крови, широко распространенная
в некоторых странах Азии и Африки. В
результате наследственного
дефекта в молекуле гемоглобина эритроциты
принимают форму
серпа и не способны выполнять свою
функцию — переносить кислород. Люди,
гомозиготные по рецессивному аллелю
серповидноклеточности
(ss),
гибнут
в возрасте 14—18 лет. Несмотря на
это частота данного аллеля достигает
в ряде районов земного шара от 8 до 20%.
При этом высокая концентрация летального
аллеля (s)
наблюдается
только в районах, где распространена
особая форма малярии, вызывающая большую
смертность населения. Случайно ли
такое совпадение? Оказалось, что
естественный отбор покровительствует
особям гетерозиготным по гену
серповидноклеточности (Ss).
Гетерозиготы
(Ss)
более
устойчивы к малярии по сравнению с
гомозиготами (SS)
по нормальному
аллелю, у которых высокая смертность
от малярии. Гомозиготы по рецессивному
аллелю (ss)
хотя и
устойчивы к малярии, но гибнут от
серповидноклеточной анемии. Так сложное
разнонаправленное
действие отбора на устойчивость к
малярии и на устранение аллеля
серповидноклеточности приводит к
существованию в состоянии длительного
равновесия двух генетически различных
форм — гомо-
и гетерозигот по серповидноклеточной
анемии. Это явление
носит название сбалансированного
полиморфизма. Таким
образом, взаимопроникновение эволюционной
теории и популяционной генетики позволило
более точно изучить действие естественного
отбора в популяции и моделировать многие
ситуации, имеющие место в процессе
эволюции.
Отбор
в пользу гетерозигот. Нередко
наблюдается отбор в пользу
гетерозигот, когда обе гомозиготы имеют
пониженную приспособленность
по сравнению с гетерозиготами. Хорошо
известным примером
такого отбора в популяциях человека
является серповидноклеточная анемия
— болезнь крови, широко распространенная
в некоторых странах Азии и Африки. В
результате наследственного
дефекта в молекуле гемоглобина эритроциты
принимают форму
серпа и не способны выполнять свою
функцию — переносить кислород. Люди,
гомозиготные по рецессивному аллелю
серповидноклеточности
(ss),
гибнут
в возрасте 14—18 лет. Несмотря на
это частота данного аллеля достигает
в ряде районов земного шара от 8 до 20%.
При этом высокая концентрация летального
аллеля (s)
наблюдается
только в районах, где распространена
особая форма малярии, вызывающая большую
смертность населения. Случайно ли
такое совпадение? Оказалось, что
естественный отбор покровительствует
особям гетерозиготным по гену
серповидноклеточности (Ss).
Гетерозиготы
(Ss)
более
устойчивы к малярии по сравнению с
гомозиготами (SS)
по нормальному
аллелю, у которых высокая смертность
от малярии. Гомозиготы по рецессивному
аллелю (ss)
хотя и
устойчивы к малярии, но гибнут от
серповидноклеточной анемии. Так сложное
разнонаправленное
действие отбора на устойчивость к
малярии и на устранение аллеля
серповидноклеточности приводит к
существованию в состоянии длительного
равновесия двух генетически различных
форм — гомо-
и гетерозигот по серповидноклеточной
анемии. Это явление
носит название сбалансированного
полиморфизма. Таким
образом, взаимопроникновение эволюционной
теории и популяционной генетики позволило
более точно изучить действие естественного
отбора в популяции и моделировать многие
ситуации, имеющие место в процессе
эволюции.
Формы естественного отбора в популяциях Интенсивность давления отбора — его количественная характеристика, направление естественного отбора определяет качественное его влияние на эволюцию. В зависимости от направления выделяют разные формы естественного отбора.
Генетической основой любой формы естественного отбора является наследственная изменчивость, а причиной — влияние условий среды. Мутанты, бывшие прежде менее приспособленными по сравнению с нормальным генотипом, при благоприятном для них изменении условий среды получают преимущество и постепенно вытесняют прежнюю норму. Результатом длительного действия отбора является преобразование популяционного генофонда, замена одних, количественно преобладающих, генотипов другими. В синтетической теории эволюции выделяли три формы естественного отбора. В настоящее время их известно 4.
 Движущая
форма естественного отбора. Движущий
отбор был описан
еще Ч. Дарвином. Само название «движущий»
говорит о том,
что такой отбор выступает в качестве
творческой силы эволюции.
При движущей форме отбора происходит
отсев мутаций с одним значением среднего
признака, которые заменяются мутациями
с другим средним значением признака.
Эта форма отбора выявляется легче
других. В итоге действия движущей формы
отбора, например, возникает увеличение
размеров потомков по сравнению
с предками (в эволюционном ряду лошадиных
от имевшего размеры
с лисицу ископаемого фенакодуса до
современных осла, зебры,
лошади). Другие формы могут уменьшаться
в размерах. Так,
на острова Средиземного моря попали в
конце третичного периода
слоны. В условиях ограниченных ресурсов
островных лесов
Движущая
форма естественного отбора. Движущий
отбор был описан
еще Ч. Дарвином. Само название «движущий»
говорит о том,
что такой отбор выступает в качестве
творческой силы эволюции.
При движущей форме отбора происходит
отсев мутаций с одним значением среднего
признака, которые заменяются мутациями
с другим средним значением признака.
Эта форма отбора выявляется легче
других. В итоге действия движущей формы
отбора, например, возникает увеличение
размеров потомков по сравнению
с предками (в эволюционном ряду лошадиных
от имевшего размеры
с лисицу ископаемого фенакодуса до
современных осла, зебры,
лошади). Другие формы могут уменьшаться
в размерах. Так,
на острова Средиземного моря попали в
конце третичного периода
слоны. В условиях ограниченных ресурсов
островных лесов
преимущество имели особи с небольшими размерами. Мутации карликовости подхватывались движущей формой отбора, а исходные аллели, определявшие нормальный для слонов размер, отсеивались вследствие гибели крупных особей. В результате на островах Средиземноморья возникли карликовые слоны ростом до полу тора метров (они были истреблены первыми охотниками, заселившими эти острова). Ч. Дарвин объяснял происхождение многих бескрылых насекомых, живущих на океанических островах, действием движущего отбора.
Классическим примером действия движущего отбора в природе служит так называемый индустриальный меланизм. В районах, не подвергавшихся индустриализации, у бабочки березовой пяденицы белая окраска соответствует светлой березовой коре. Среди светлых бабочек на стволах берез встречались и темные, но они были хорошо заметны и склевывались птицами. Развитие промышленности привело к загрязнению воздуха, и белые березы покрылись слоем копоти. Теперь на темных стволах птицы гораздо легче замечали не темных, а типичных светлых бабочек Постепенно в загрязненных районах частота встречаемости темных (мутантных) особей резко возросла и они стали преобладающими, хотя еще сравнительно недавно встречались исключительно редко. Убедительный пример движущего отбора — выработка у микроорганизмов, насекомых, мышевидных грызунов устойчивости к антибиотикам и ядохимикатам. Многочисленными исследованиями установлено, что воздействие на микроорганизмы различными антибиотиками обусловливает за относительно короткий срок устойчивость к дозам, во много раз превышающим исходную Это объясняется тем, что антибиотики выступают в качестве фактора отбора, способствующего выживанию устойчивых к нему мутантных форм. Благодаря быстрому размножению микроорганизмов, мутантные особи увеличиваются в числе и образуют новые популяции, невосприимчивые к действию антибиотиков. Увеличение дозы или применение более сильных препаратов вновь создает условия для действия движущего отбора, в результате которого образуются все более и более устойчивые популяции микроорганизмов. Вот почему в медицине неуклонно идет поиск новых форм антибиотиков, к которым еще не приобрели устойчивость патогенные микробы.
В странах с передовой сельскохозяйственной культурой все чаще отказываются от химических средств защиты растений от вредителей (насекомых, грибков). Поскольку через ограниченное число поколений движущим отбором фиксируются у вредителей мутации устойчивости к химическим веществам. Вместо химической обработки признано целесообразным через 10—12 лет заменять старый сорт новым, которого еще «не нашли» вредители.
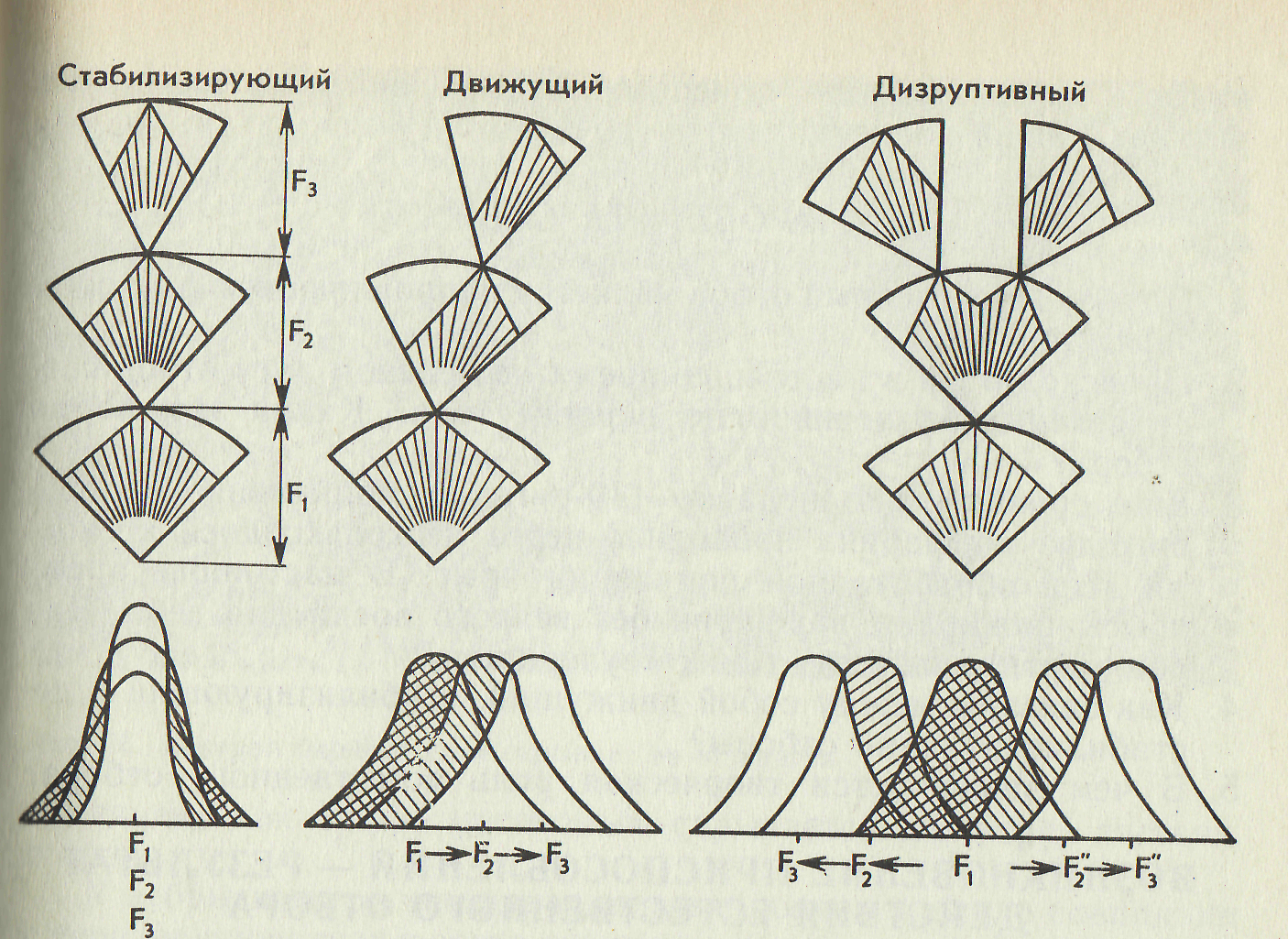
Рис. Схема действия разных форм отбора.
Стабилизирующий отбор. Известно, что реликтовое растение гинкго и потомок первоящеров гаттерия, а также кистеперая рыба — латимерия существуют почти без изменения миллионы лет.
Как объяснить такую стабильность видов, если в природе постоянно совершается мутационный процесс? Ответ на этот вопрос дает учение о стабилизирующем отборе, разработанное крупнейшим эволюционистом И. И. Шмальгаузеном.
Стабилизирующий отбор наблюдается в том случае, если условия внешней среды длительное время остаются достаточно постоянными. В относительно неизменной среде преимуществом обладают типичные, хорошо приспособленные к ней особи со средним выражением признака, а отличающиеся от них мутанты погибают. Известно много примеров стабилизирующего отбора. Так, после снегопада и сильных ветров в Северной Америке было найдено 136 оглушенных, полуживых домовых воробьев, 72 из них выжили, а 64 погибли. Погибшие птицы имели или очень длинные, или очень короткие крылья. Особи со средними, «нормальными», крыльями оказались более выносливыми.
 Стабилизирующий
отбор ведет к большой фенотипической
однородности популяции. Если он
действует длительное время, то создается
впечатление, что популяция или вид не
изменяются. Однако
эта неизменность кажущаяся и касается
лишь внешнего облика популяции,
генофонд же ее продолжает изменяться
на основе появления мутаций с тем же
значением средней, но с более узкой
нормой реакции.
Стабилизирующий
отбор ведет к большой фенотипической
однородности популяции. Если он
действует длительное время, то создается
впечатление, что популяция или вид не
изменяются. Однако
эта неизменность кажущаяся и касается
лишь внешнего облика популяции,
генофонд же ее продолжает изменяться
на основе появления мутаций с тем же
значением средней, но с более узкой
нормой реакции.
Стабилизирующая форма отбора характерна и для человека.
Известно, что нарушения по самым мелким 21—22-й парам хромосом ведут к тягчайшему наследственному заболеванию — синдрому Дауна. Если возникнут отклонения в числе и форме более крупных хромосом, это приведет к гибели оплодотворенных яйцеклеток. Самопроизвольные (спонтанные) аборты часто вызваны гибелью эмбрионов с ненормальностями в хромосомах средних размеров.
Таким образом, стабилизирующая форма отбора в течение сотен тысяч и миллионов поколений оберегает виды от существенных изменений, от разрушающего влияния мутационного процесса, выбраковывая мутантные формы. Без стабилизирующего отбора не было бы устойчивости (стабильности) в живой природе.
Стабилизирующий и движущий отборы взаимосвязаны и представляют две стороны одного процесса. Популяции постоянно вынуждены приспосабливаться к изменениям условий среды. Движущий отбор будет сохранять генотипы, которые наиболее соответствуют изменениям среды. Когда условия среды стабилизируются, отбор приведет к созданию хорошо приспособленной к ней формы. С этого момента в действие вступает стабилизирующий отбор, который будет поддерживать типичные, преобладающие генотипы и устранять от размножения уклоняющиеся от средней нормы мутантные формы.
Дестабилизирующий отбор. Стабилизирующий отбор сужает норму реакции. Однако в природе нередки случаи, когда экологическая ниша вида со временем может оказаться более широкой. В этом случае селективное преимущество получают особи и популяции с более широкой нормой реакции, сохраняющие вместе с тем тоже среднее значение признака. В итоге идет процесс, обратный стабилизирующему отбору: преимущество получают мутации с бо лее широкой нормой реакции. Так, популяции озерных лягушек, живущие в прудах с разнородной освещенностью, с чередованием участков, заросших ряской, тростником, рогозом, с «окнами» открытой воды, характеризуются широким диапазоном изменчивости окраски (итог дестабилизирующего отбора). Наоборот, в водоемах с однородной освещенностью и окраской (пруды, сплошь заросшие ряской, или открытые пруды) диапазон изменчивости окраски лягушек узок (итог действия стабилизирующего отбора). Таким образом, дестабилизирующая форма отбора ведет к расширению нормы реакции.
Разрывающий (дизруптивный) отбор. Для многих популяций характерен полиморфизм — существование двух или нескольких форм по тому или иному признаку. Полиморфизм нельзя объяснить только возникновением новых мутаций. Причины его могут быть разными. В частности, он может быть обусловлен повышенной относительной жизнеспособностью гетерозигот. В других случаях полиморфизм может быть результатом действия особой формы отбора, получившей название разрывающего или дизруптивного. Эта форма отбора осуществляется в тех случаях, когда две или более генетически различные формы обладают преимуществом в разных условиях, например, в разные сезоны года. Хорошо известен случай преимущественного выживания в зимнее время «красных», а в летнее – «черных» форм двуточеченой божьей коровки. Дизруптивный отбор направлен против средних промежуточных форм. Он как бы разрывает популяцию по данному признаку на несколько групп и может привести к разделению ее на две и более.
Творческая роль отбора. На первый взгляд может показаться, что естественный отбор выполняет в эволюции роль своеобразного «сита», устраняющего из популяции неблагоприятные изменения. Такая роль у отбора действительно существует, но этим не ограничивается его значение в эволюции. Через отбор признаков определяется направление эволюционного процесса, способствует возникновению приспособленности организмов к разнообразным местам обитания.
 Таким
образом, можно отметить приспособительное,
формообразовательное и интегрирующее
действие отбора на организмы.
Таким
образом, можно отметить приспособительное,
формообразовательное и интегрирующее
действие отбора на организмы.
Подводя итог сегодняшней теме, надо отметить, что наличие мутаций и естественного отбора необходимо и достаточно для приспособительной и дивиргентной эволюции организмов. Поэтому данные факторы можно обозначить, как необходимые факторы эволюционного процесса. При этом отбор – это единственный движущий и направляющий фактор, оказывающий влияние и на саму мутационную изменчивость. Остальные рассмотренные факторы (популяционные волны, изоляция, дрейф генов) – являются дополнительными факторами. Для понимания эволюционного процесса необходимо учитывать сложные взаимодействия всех факторов.
Лекция 10
Видообразование и межвидовые отношения.
План.
Аллопатрическое видообразование
Симпатрическое видообразование
Темпы видообразования.
Взаимоотношения близких видов.
Межвидовые отношения в биоценозах.
Элиминация и ее формы
Среди различных результатов эволюционного процесса видообразование в наибольшей мере привлекало внимание исследователей на всех этапах развития эволюционной теории (не случайно Ч.Дарвин назвал свой основной труд «Происхождение видов»). Именно этот аспект эволюционного процесса стал основным содержанием синтетической теории эволюции, которая, в сущности, представляет собой главным образом теорию видообразования.
Аллопатрическое видообразование
Как мы уже упоминали, всякая территориально изолированная популяция характеризуется собственным генофондом, частота встречаемости разных аллелей в котором оптимизирована отбором применительно к условиям обитания именно данной популяции. При обособлении популяции обмен наследственной информацией («поток генов») между нею и остальными популяциями прекращается. Генофонд изолированной популяции становится самостоятельным. Особенности местных условий определяют специфическое направление отбора, действующего на такую популяцию (это направление является одним из направлений дизруптивного отбора, действующего на вид в целом). Постепенно в генофонде изолированной популяции происходит накопление новых мутаций. Поскольку генофонд данной популяции специфичен, спектр его изменчивости должен несколько отличаться от такового других популяций вида. Специфика местных условий и соответствующее направление отбора усугубляют особенности генофонда изолированной популяции. Поэтому мутации, накапливающиеся в ее генофонде, будут в той или иной степени отличаться от мутаций, которые за это же время закрепятся в других популяциях родительского вида.
Если географическая изоляция достаточно продолжительна, а направления отбора достаточно различны, то изолированная популяция приобретает отчетливые различия с родительским видом, снижающие возможность скрещивания, вплоть до возникновения репродуктивной изоляции (т. е. нескрещиваемости в природных условиях) между ними. Репродуктивная изоляция может основываться на самых разных биологических особенностях близких форм организмов. Выделяют следующие основные формы репродуктивной изоляции:
1) этологическая (различия поведения, особенно брачного).
2) экологическая (различия предпочитаемых местообитаний);
3) временная, или сезонная (различия сроков размножения);
4) морфологическая (различия размеров тела и особенностей строения, важных для опознания половых партнеров, их взаимной стимуляции и спаривания);
5) генетическая (различия генома, приводящие к генетической несовместимости половых клеток).
Все эти и другие формы изоляции возникают независимо друг от друга и могут сочетаться в любых комбинациях или действовать поодиночке. Наиболее совершенна генетическая изоляция, исключающая всякую возможность обмена генетической информацией между двумя популяциями.
После возникновения той или иной формы репродуктивной изоляции обособленная популяция становится самостоятельным видом. Если теперь географический барьер, разделявший популяции, нарушится и вновь восстановится контакт между ними — эти два вида либо смогут сосуществовать в одном ареале, либо между ними возникнут конкурентные отношения и один из них будет вытеснять другой, если их биология осталась близкой. Но, так или иначе, это будут уже две обособленные филетические линии, каждая из которых имеет свою эволюционную судьбу.
Такая форма видообразования, основанная на территориальной изоляции разных популяций, получила название аллопатрической. Ч.Дарвин также указывал, что географическая изоляция способствует видообразованию, но придавал этому фактору меньшее значение, чем современная теория эволюции.
Если две изолированные друг от друга популяции одного вида находятся в сходных условиях, длительное время остающихся постоянными, на них будет действовать одинаково направленный стабилизирующий отбор. В таких популяциях формируются практически одинаковые оптимальные фенотипы. Однако под прикрытием этих оптимальных фенотипов в генофонде популяций, как мы видели выше, происходит постепенное накопление новых мутаций. Если обмен генетической информацией между двумя такими популяциями затруднен в течение достаточно длительного времени, в их генофондах могут накапливаться разные мутации. В результате популяции, оставаясь внешне сходными, будут все более различаться в генетическом отношении. С накоплением этих различий возможности их гибридизации постепенно снижаются, вплоть до полной невозможности скрещивания (возникновение генетической несовместимости) особей из разных популяций. На этой стадии две обособленные популяции превращаются в два самостоятельных вида, которые, однако, сохраняют фенотипическое сходство. Такие виды носят название видов-двойников. Сходство видов-двойников может быть столь полным, что даже опытные зоологи-систематики нередко с большим трудом различают подобные формы, единственным надежным критерием для разделения которых является их нескрещиваемость друг с другом.
 Изолированные
популяции могут возникать двумя путями:
либо при появлении каких-либо географических
барьеров, разделяющих прежний ареал
вида, либо при активном расселении вида
на новые территории за пределы занимаемого
им ареала. В этом случае относительно
небольшие популяции, укоренившиеся
в новых районах, могут с самого момента
своего возникновения стать
территориально обособленными
(периферические
изоляты). Этот
последний случай особенно интересен
тем, что периферические изоляты могут
сразу подвергнуться действию условий,
существенно отличающихся от таковых в
пределах ареала вида. Э.
Майр высказал
предположение, что в периферических
изолятах могут в некоторых случаях
происходить относительно очень быстрые
и резкие изменения генофонда, приводящие
к эволюционным изменениям значительного
масштаба. Этот процесс
получил название «генетическая
революция». Основную
причину
генетической революции Э.Майр видит в
том, что генофонд периферического
изолята (или популяции
основателей) с
самого начала не сбалансирован и не
оптимизирован отбором. Поэтому при
закреплении популяции основателей в
новом для вида районе ее генофонд должен
подвергнуться существенной перестройке
под давлением отбора, до установления
новой формы генетического равновесия.
При малочисленности популяций основателей
преобразования их генофондов ускоряются
явлениями дрейфа генов.
Изолированные
популяции могут возникать двумя путями:
либо при появлении каких-либо географических
барьеров, разделяющих прежний ареал
вида, либо при активном расселении вида
на новые территории за пределы занимаемого
им ареала. В этом случае относительно
небольшие популяции, укоренившиеся
в новых районах, могут с самого момента
своего возникновения стать
территориально обособленными
(периферические
изоляты). Этот
последний случай особенно интересен
тем, что периферические изоляты могут
сразу подвергнуться действию условий,
существенно отличающихся от таковых в
пределах ареала вида. Э.
Майр высказал
предположение, что в периферических
изолятах могут в некоторых случаях
происходить относительно очень быстрые
и резкие изменения генофонда, приводящие
к эволюционным изменениям значительного
масштаба. Этот процесс
получил название «генетическая
революция». Основную
причину
генетической революции Э.Майр видит в
том, что генофонд периферического
изолята (или популяции
основателей) с
самого начала не сбалансирован и не
оптимизирован отбором. Поэтому при
закреплении популяции основателей в
новом для вида районе ее генофонд должен
подвергнуться существенной перестройке
под давлением отбора, до установления
новой формы генетического равновесия.
При малочисленности популяций основателей
преобразования их генофондов ускоряются
явлениями дрейфа генов.
Г.Льюис подкрепил гипотезу Э.Майра о возможности генетических революций анализом последствий действия на такие популяции особенно интенсивного отбора, названного им «катастрофическим». Популяции, обитающие в краевой зоне ареала вида, часто подвергаются действию необычных для данного вида экстремальных условий. При крайне неблагоприятных условиях на популяцию действует катастрофический отбор, элиминирующий большинство составляющих ее особей. При этом выживают немногие индивиды, обладающие наиболее высокой фенотипической устойчивостью к неблагоприятным факторам (на основе соответствующих генетических уклонений). После возвращения условий к норме численность популяции восстанавливается, но се генофонд после действия катастрофического отбора должен существенно отличаться от исходного.