

“УТВЕРЖДАЮ’’
Зав.кафедры биологии
Д.м.н. М.В. Черников
“19” сентября 2011г.
М Е Т О Д И Ч Е С К И Е У К А З А Н И Я
к практическим занятиям по биологии для преподавателей.
ТЕМА: Размножение организмов. Бесполое и половое. Гаметогенез. Мейоз.
ЦЕЛЬ ЗАНЯТИЯ:
Получить представление о размножении как одном их основных критериев жизни. Знать основные формы размножения и кардинальные отличия бесполого и полового размножения. Знать особенности ово- и сперматогенеза и суть основных этапов и фаз мейоза, а также качественные изменения происходящие в хромосомах в процессе двух делений.
Место проведения: учебная аудитория кафедры биологии
Время проведения 2 часа
ПЕРЕЧЕНЬ ЗНАНИЙ И ПРАКТИЧЕСКИХ НАВЫКОВ:
1. Знать особенности бесполого размножения и его биологическое значение.
2. Знать особенности полового размножения и его биологическую сущность.
3. Уметь находить на микропрепаратах отличия между половыми клетками.
4. Знать гаметогенез. Цитологические основы этапов сперматогенеза и овогенеза.
5.Знать особенности оплодотворение и его биологическая сущность.
6. Партеногенез, и его виды.
7. Знать характеристику каждой фазы мейоза.
8. Знать особенности перестройки хромосом и перекомбинации генов в процессе двух мейотических делений.
9. Знать механизм и биологический смысл кроссинговера.
10.Знать различия мейоза у низших и высших организмов.
ОСНОВНЫЕ ВОПРОСЫ ТЕМЫ:
1. Бесполое размножение у одноклеточных и многоклеточных организмов.
2. Половое размножение и его биологическое значение.
3. Особенности строения половых клеток. Гаметогенез. Цитологические основы этапов сперматогенеза и овогенеза.
4. Партеногенез, и его виды.
7. Знать характеристику каждой фазы мейоза.
8. Знать особенности перестройки хромосом и перекомбинации генов в процессе двух мейотических делений.
9. Знать механизм и биологический смысл кроссинговера.
10.Знать различия мейоза у низших и высших организмов.
Технологическая карта занятия.
№ |
Этап занятия |
Время в минутах |
1 |
Организация занятия |
2 |
2 |
Определение темы и цели занятия |
3 |
3 |
Определение Исходного уровня знаний |
20 |
4 |
Самостоятельная работа студентов |
90 |
5 |
Подведение итогов занятия, проверка протоколов |
5 |
Учебные пособия |
|
Средства обучения |
Таблицы, микропрепараты |
Методические указания Практикумы Чебышев Н.В. и др. «Руководство к лабораторным занятиям по биологии» 2008г. Руководство к лабораторным занятиям по биологии. /Под ред. Ю.Д. Богоявленского. - М.: Медицина, 1998.
|
|
СОДЕРЖАНИЕ САМОСТОЯТЕЛЬНОЙ РАБОТЫ:
КРАТКОЕ СОДЕРЖАНИЕ ТЕМЫ:
Воспроизведение себе подобных у человека является уникальным процессом, в основе которого лежит образование половых клеток - гаметогенез. Гаметогенез обеспечивает генетическую приемственность в приобретении организмами биологической информации. Нарушение процессов гаметогенеза приводит к образованию атипичных зигот, которые реализуются либо в ранние недиагностируемые выкидыши, либо являются причиной различных патологий у новорождённых.
Размножение – универсальное свойство живых организмов, обеспечивающее материальную непрерывность в ряду поколений. Существует два основных способа размножения: бесполое и половое. При бесполом размножении начало новому организму дают соматические клетки (митотическое деление, шизогония, почкование, спорообразование, вегетативное размножение). Половое размножение совершается с участием половых клеток, гамет. Особенностью половых клеток является гаплоидный набор хромосом.
Оплодотворение – соединение двух гамет, в результате чего образуется зигота. Процессу оплодотворения предшествует сближение мужской и женской половых клеток, т.е. объединение наследственного материала материнского и отцовского организмов.
Партеногенез. Особую форму полового размножения представляет собой партеногенез (гр. рагthenos — девственница, genos— рождение) т. е. развитие организма из неоплодотворенных яйцевых клеток. Эта форма размножения была обнаружена в середине XVIII в. швейцарским натуралистом Ш. Бонне (1720—1793). В настоящее время известен не только естественный, но и искусственный партеногенез.
Естественный партеногенез существует у ряда растений, червей, насекомых, ракообразных. У некоторых животных любое яйцо способно развиваться как без оплодотворения, так и после него. Это так называемый факультативный (необязательный) партеногенез. Он встречается у пчел, муравьев, коловраток, у которых из оплодотворенных яиц развиваются самки, а из неоплодотворенных — самцы. У этих животных партеногенез возник как приспособление для регулирования численного соотношения полов.
При облигатном, т. е. обязательном, партеногенезе яйца развиваются без оплодотворения. Этот вид партеногенеза известен, например, у кавказской скальной ящерицы. У многих видов партеногенез носит циклический характер. У тлей, дафний, коловраток в летнее время существуют лишь самки, размножающиеся партеногенетически, а осенью партеногенез сменяется размножением с оплодотворением (это явление получило название гете- рогении;). Облигатный и циклический партеногенез исторически развивался у тех видов животных, которые погибали в большом количестве (тли, дафнии) или у которых была затруднена встреча особей различного пола (скальные ящерицы). Вид кавказской скальной ящерицы сохранился лишь благодаря появлению партеногенеза, так как встреча двух особей, обитающих на скалах, отделенных глубокими ущельями, затруднена. В настоящее время все особи этого вида представлены лишь самками, размножающимися партеногенетнчески.
В ядрах соматических клеток особей, развившихся из неоплодотворенных яиц, в ряде случаев имеется гаплоидный набор хромосом (таковы самки коловраток), в других — диплоидный (тли, дафнии).
Искусственный партеногенез исследовал А.А. Тихомиров. Он добился развития неоплодотворенных яиц тутового шелкопряда, раздражая их тонкой кисточкой или обрабатывая в течение нескольких секунд серной кислотой.
Тот факт, что дробление яйца начинается только после его оплодотворения, получил объяснение благодаря опытам с искусственным партеногенезом. Опыты показали, что для развития яйца необходима активация, которая является следствием тех сдвигов в обмене веществ, которые сопутствуют оплодотворению. В естественных условиях эти сдвиги происходят после проникновения сперматозоида в яйцеклетку, но в эксперименте могут быть вызваны разнообразными воздействиями: химическими, механическими, электрическими, термическими и др. проникновение сперматозоида, влекут за собой обратимые повреждения протоплазмы яйцеклетки, что изменяет метаболизм и оказывает активирующее воздействие.
Гаметогенез – формирование яйцеклеток и сперматозоидов в организме. Этот процесс
происходит в половых железах – гонадах, которые представлены семенниками у
самцов и яичниками у самок. Процесс развития мужских половых клеток называется сперматогенезом (sperma – семя, genesis – происхождение), женских половых клеток – овогенезом (ovum - яйцо).
Сперматогенез – это процесс превращения исходно диплоидных стволовых клеток в гаплоидные сперматозоиды. В нем выделяются 4 периода: 1.период размножения; 2.период роста;
3.Период созревания; 4.Период формирования сперматозоида (спермиогенез).
Сперматогенез происходит в мужских гонадах – семенниках (testis). В течение периода размножения диплоидные мужские половые клетки – сперматогонии - делятся путем митоза несколько раз. Последующие генерации сперматогоний отличаются от первых по величине и степени конденсации хроматина.
Второй период сперматогенеза – период роста. Половые клетки на этой стадии называются сперматоцитами первого порядка (сперматоциты 1). Они несколько увеличиваются в объеме, а в их ядрах происходит профаза мейоза, во время которой осуществляются подготовка к редукции числа хромосом, конъюгация и кроссинговер. Последовательные события профазы мейоза происходят в ходе стадий лептотены, зиготены, пахитены и диплотены.
В течение третьего периода, периода созревания, совершаются два деления мейоза и осуществляется редукция числа хромосом. При первом делении (редукционном) к полюсам расходятся гомологичные хромосомы, при втором делении (эквационном) – сестринские хроматиды. Второе деление созревания быстро следует за первым, иногда без собственной интерфазы. После первого деления созревания клетки сперматогенного ряда называются сперматоцитами второго порядка (сперматоцитами II), а после второго деления созревания – сперматидами. Таким образом, из каждого сперматоцита I получается два сперматоцита II, а затем четыре одинаковые по размерам гаплоидные клетки – сперматиды. Во время третьего периода сперматогенеза происходит: рекомбинация генов в гомологичных хромосомах ( кроссинговер) и образуются гаплоидные клетки.
Процесс развития женских половых клеток – овогенез – имеет ряд общих черт с процессом сперматогенеза и целый ряд отличий. Оогенез совершается в три этапа, называемых периодами:
Период размножения; 2.Период роста; 3.Период созревания.
В период размножения диплоидные оогонии совершают определенное, характерное для
данного вида число митотических делений, а затем переходят к стадии роста и становятся ооцитами первого порядка(ооцитI). Особенностью оогенеза, отличающей его от сперматогенеза, является меньшая митотическая активность оогониев. У высших позвоночных пролиферация оогоний происходит только в эмбриональный период и запас половых клеток (ооцитов) в яичниках взрослых самок не пополняется.
В период роста осуществляется одновременно два процесса: в ядре происходит профаза мейоза I, а в цитоплазме растущего ооцита накапливаются органеллы, питательные вещества, необходимые для обеспечения потребностей зародыша. Рост овоцита неравномерен. Важным событием является редупликация ДНК.
Период созревания – заключительный этап оогенеза – осуществляется в виде двух последовательных делений созревания. В результате первого деления мейоза гомологичные хромосомы расходятся в дочерние ядра, в результате второго деления созревания в клетках оказывается гаплоидное число хромосом. Особенностью делений созреванья в оогенезе является асимметричное разделение цитоплазмы: при каждом делении почти вся цитоплазма остается в одной из сестринских клеток, тогда как в другой её объем ничтожен. При первом делении образуются большой ооцит второго порядка (ооцит II) и маленькое первое редукционное тельце. В цитоплазме ооцита содержится практически все, что накоплено в период роста. При втором делении созревания образуется зрелое яйцо с тремя редукционными тельцами, которые в последствии обычно дегенерируют.
Женская половая клетка – яйцеклетка – высокоспециализированная гаплоидная клетка организма, обладающая потенциальной способностью к развитию. В её цитоплазме запасено всё, что необходимо для роста и развития зародыша. Ооцит активно накапливает различные трофические компоненты и органеллы, РНК, структурные белки, а также желток, который служит источником питания для зародыша. Количество и распределение желтка имеют важное значение для развития зародыша. Кроме плазматической оболочки, характерной для каждой живой клетки, яйцеклетки обычно окружены специализированными оболочками. Различают первичные оболочки (желточные), которые формируются самим яйцом, вторичные оболочки – образуются фолликулярными клетками, окружающими половую клетку, и третичные оболочки – выделяются клетками яйцевода при прохождении по нему овулировавшего яйца.
Мейозом называется особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в 2 раза (от древнегреч. «мейон» - меньше - и от «мейозис» - уменьшение). Исходное число хромосом в мейоцитах (клетках, вступающих в мейоз) называется диплоидным хромосомным числом (2n) Число хромосом в клетках, образовавшихся в результате мейоза, называется гаплоидным хромосомным числом (n). Главной особенностью мейоза является конъюгация (спаривание) гомологичных хромосом с последующим расхождением их в разные клетки. Мейотическое распределение хромосом по дочерним клеткам называется сегрегацией хромосом.
Динамика мейоза
Типичный мейоз состоит из двух последовательных клеточных делений, которые соответ- ственно называются мейоз 1 и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным, реже ге- гетеротипным. Во втором делении число хромосом не изменяется, такое деление называют экваиионным (уравнивающим), реже - гомеотипным. Выражения «мейоз» и «редукционное деление» часто используют как синонимы.
В ходе двух делений мейоза ДНК делится поровну между 4 клетками. Мейоз длится много дольше митоза: например, у пшеницы он продолжается 24 часа, у лилии — 9—12 суток, у мыши — 11—14 суток, у человека (мужчины) — 24 дня. У ряда животных и человека во время образования женских половых клеток мейоз останавливается на срок до нескольких лет и завершается только во время оплодотворения.
Интерфаза
Предмейотическая интерфаза отличается от обычной интерфазы тем. что процесс репликации ДНК не доходит до конца: примерно 0,2.0,4 % ДНК остается неудвоенной. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с. При наличии центриолей происходит их удвоение таким образом, что в клетке имеется две диплосомы, каждая из которых содержит пару центриолей.
Первое деление мейоза (редукционное деление, или мейоз I). Сущность редукционного деления заключается в уменьшении числа хромосом в два раза: из исходной диплоидной клетки образуется две гаплоидные клетки с двухроматидными хромосомами (в состав каждой хромосомы входит 2 хроматиды. n2с).
Профаза 1. (профаза первого деления) состоит из ряда стадий: лептонемы,зигонемы. пахинемы. диплонемы, диктиотены и диакинеза.
• Лептонема (стадия тонких нитей). Гомологичные хромосомы начинают коньюгировать в одних участках, оставаяс разобщенными в других. Лептонемные хромосомы отличаются от хромосом обычной профазы митоза в двух отношениях: они «выглядят» в продольном направлении едиными, а не двойными, хотя синтез ДНК уже прошел, и следовательно они двойные. По всей длине с неравномерными промежутками располагаются плотные утолщения-хромомеры. Хромомеры представляют собой участки хроматина, уплотнившиеся в результате локального сокращения хроматинового вещества. Хромосомы видны в световой микроскоп в виде клубка тонких нитей.
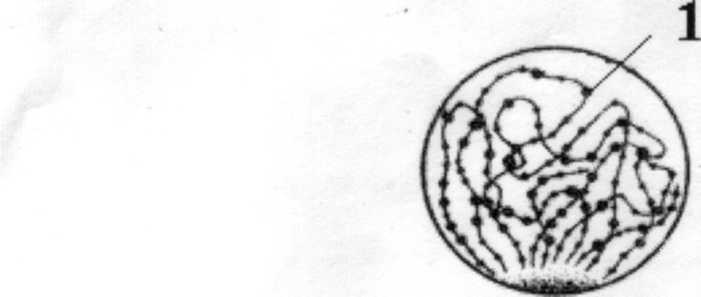
Рисунок 1. Стадия лептонемы. 1- хромомеры.
• Зигонема (стадия сливающихся нитей). Происходит конъюгация гомологичных хромосом (от лат. conjugatio - соединение, спаривание, временное слияние). При конъюгации образуются биваленты. Каждый бивалент - это относительно устойчивый комплекс из одной пары гомологичных хромосом. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов.
Объединение гомологов начинается на концах хромосом - в теломерах, а гак же в центромерных районах, позднее объединение происходит внутри бивалента.
Количество бивалентов равно гаплоидному числу хромосом. Иначе биваленты называются тетрады, так как в состав каждого бивалента входит 4 хроматиды.

Пахинема (стадия толстых нитей). Хромосомы спирализуются, хорошо видна их продольная неоднородность.Завершается репликация ДНК (образуется особая пахинемная ДНК). Хромосомы несколько укорачиваются и утолщаются. Между хроматидами материнского и отцовского происхождения в нескольких местах возникают соединения - хиазмы (греч. chiasma - перекрест), или рекомбинантные узелки. Они представляют собой белковые комплексы размерами около 90 нм. В области каждой хиазмы происходит обмен соответствующих участков гомологичных хромосом - от отцовской к материнской и наоборот. Этот процесс называют кроссинговером (англ. crossing-over - перекрест). Кроссинговер осуществляется с очень большой точностью, поэтому ни одна из хроматид не теряет и не приобретает генов. Более того, если кроссинговер происходит в последовательности одного гена, то не теряется и не приобретается ни один нуклеотид в хроматидах, обменивающихся участками гомологичной ДНК.
Таким образом, кроссинговер обеспечивает многочисленные генетические рекомбинации. В каждом биваленте человека в профазе 1 кроссинговер происходит в среднем в двух-трех участках. Количество рекомбинантных узелков равно количеств) перекрестков.
• Дитотена (стадия двойных нитей- от греческого диплос- двойной). Каждая хромосома состоит из двух ясно различимых хроматид. Следовательно, каждый бивалент слагается из четырёх хроматид. Однако гомологи разделяются не полностью. В одной или нескольких точках контакт между ними сохраняется, эти точки называются хиазмы. Каждая хиазма образуется в результате кроссинговера. Хиазм образуется больше в крупных хромосомах, всего же на одну гамету приходится примерно 40 кроссинговеров.
Некоторые исследователи считают, что отсутствие хиазм в биваленте является фактором, предрасполагаюшим к нерасхождению хромосом. По количеству хиазм можно судить об интенсивности кроссинговера. Если образуется только одна хиазма, то бивалент на стадии диплонемы имеет вид креста. Если образуются две хиазмы, то бивалент имеет форму кольца, при трёх или более числе хиазм у гомологов развивается ряд петель.
Поскольку сжатие и отталкивание бивалентов усиливается, хиазмы двигаются к концам хромосом, т. е. происходит терминализация хиазм. В конце диплонемы наступает деспирализация хромосом; гомологи продолжают отталкиваться друг от друга.
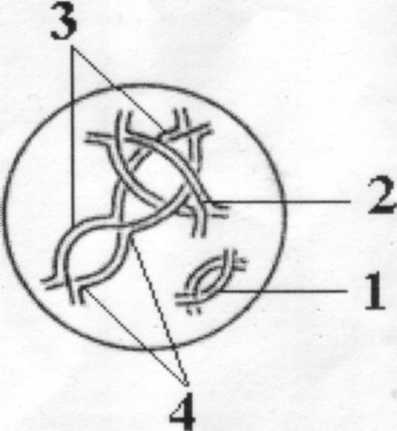
Рисунок 3. Диплотена. 1- кольцо образованное двумя хиазмами, 2 - одна хиазма образует фигуру -крест, 3 - три хиазмы образуют фигуру восьмерку, 4-хиазмы.
• Диктиотена. Стадия характерная только для овогенеза, при сперматогенезе она отсутствует. Ооциты I на этой стадии приостанавливают свое развитие, и их деление завершается после полового созревания. Хромосомы на этой стадии имеют вид «ламповых щеток». Такая структура образуется в результате деконденсации хроматина. И состоят хромосомы из длинной нити, на которой находятся хромомеры. Каждая хромомера имеет пару, от хромомер отходят петли. Поэтому такие хромосомы похожи на ершик.
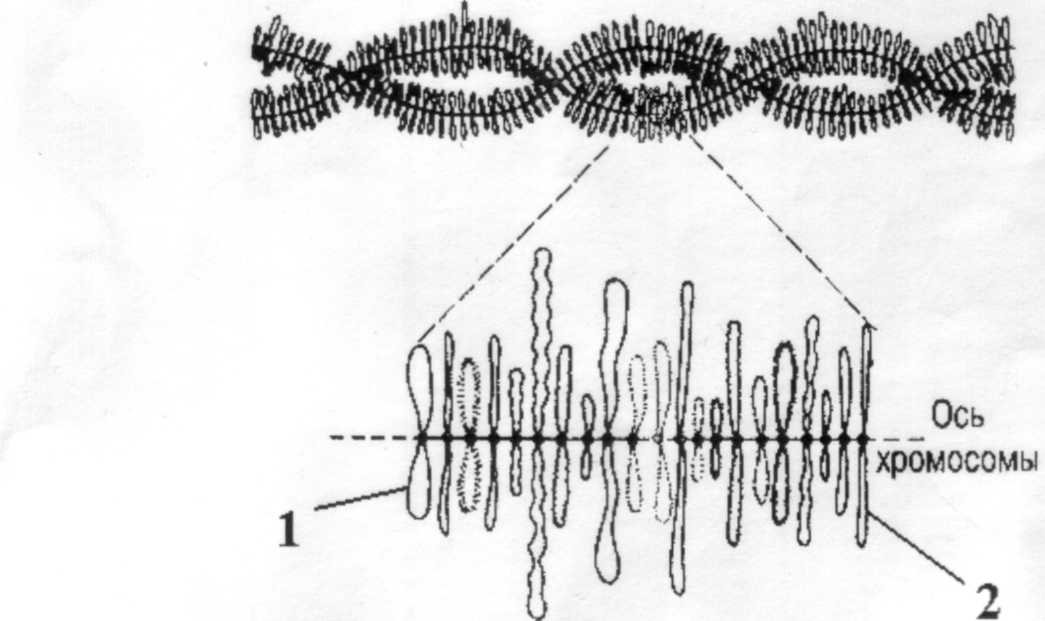
Рисунок 4. Хромосомы «типа ламповых» щеток. 1,2- неконденсированные петли.
Диакинез (стадия расхождения бивалентов). Отдельные биваленты располагаются на периферии ядра. В диакинезе продолжается конденсация хромосом, они отделяются от нуклеолеммы. но гомологичные хромосомы продолжают еще оставаться связанными между собой хиазмами, а сестринские хроматиды каждой хромосомы - центромерами. Благодаря наличию нескольких хиазм биваленты образуют петли. В это время разрушаются ядерная оболочка и ядрышки. Реплицированные центриоли направляются к полюсам, образуется веретено деления.
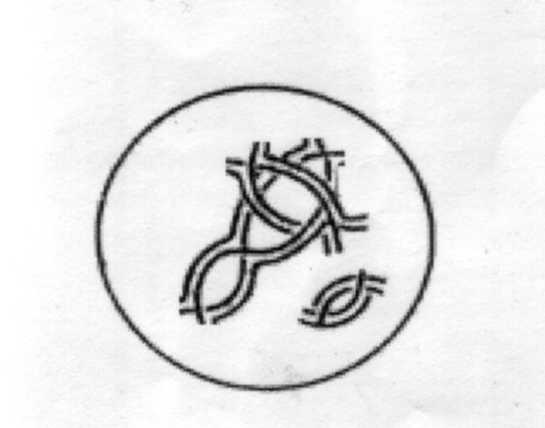
Рисунок 5. Стадия диакенеза. Хиазмы сдвинуты к теломерным участкам хромосом.
Метафаза I (метафаза первого деления).
В прометафазе I ядерная оболочка разрушается (фрагментируется). Формируется веретено деления. Далее происходит метакинез - биваленты перемещаются в
экваториальную плоскость клетки, хромосомные микротрубочки прикрепляются к центромере лишь с одной стороны (со стороны полюса), а центромеры гомологичных хромосом расположены по обеим сторонам экватора. Связь между хромосомами с помощью хиазм продолжает сохраняться. Форма каждого бивалента определяется числом и локализацией хиазм. У мужчин число хиазм на бивалент в метафазе 1 составляет обычно 1-5. Бивалент ХУ становится палочковидным в результате одиночной терминально расположенной хиазмы.

Рисунок 6. Метафаза I, биваленты располагаются в плоскости экватора. Анафаза I (анафаза первого деления).
Рисунок
7. Анафаза I. К полюсам
расходятся гомологичные хромосомы.

Телофаза I (пгелофаза первого деления).
Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. В норме каждая дочерняя клетка получает одну гомологичную хромосому из каждой пары гомологов. Формируются два гаплоидных ядра, которые содержат в два раза меньше хромосом, чем ядро исходной диплоидной клетки. Каждое гаплоидное ядро содержит только один хромосомный набор, то есть каждая хромосома представлена только одним гомологом. Содержание ДНК в дочерних клетках составляет 2с. В большинстве случаев (но не всегда) телофаза I сопровождается цитокинезом.

Рисунок 8. Телофаза I. Образование двух гаплоидных клеток.
Интеркинез
Интеркинез - это короткий промежуток между двумя мейотическими делениями. Отличается от интерфазы тем. что не происходит репликации ДНК, удвоения хромосом и удвоения центриолей: эти процессы произошли в предмейотической интерфазе и, частично, в профазе I.
Второе деление мейоза (эквационное деление, или мейоз II).
В ходе второго деления мейоза уменьшения числа хромосом не происходит. Сущность эквационного деления заключается в образовании четырех гаплоидных клеток с однохроматидными хромосомами (в состав каждой хромосомы входит одна хроматида, пс).
Профаза II (профаза второго деления).
Не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей. В каждой из дочерних клеток формируется веретено деления.
Метафаза II (метафаза второго деления).
Рисунок
9. Метафаза II. Хромосомы на
экваторе клетки.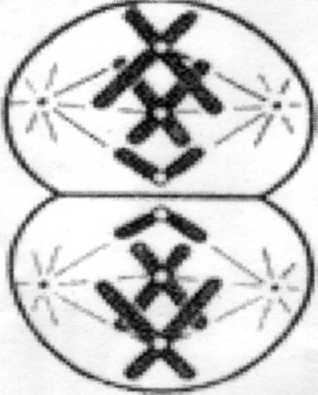
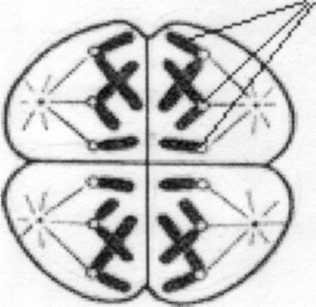
Рисунок 10. Телофаза II. Образование 4-х гаплоидных клеток. 1-сетринские хроматиды.
Отклонения от типичного мейоза.
У высших организмов отклонение от типичного мейоза обычно рассматривается как аномалия. Наиболее частым отклонением от типичного мейоза является нарушение нормальной сегрегации хромосом: их нерасхождение или в первом, или во втором делении. Нерасхождение хромосом (как и при митозе) может быть обусловлено действием внешних факторов, но может быть и спонтанным. Рассмотрим типичные нарушения сегрегации хромосом.