

Разнообразие форм днк
Существовавшее до недавнего времени мнение о том, что В-ДНК — это совершенная двойная спираль, геометрия которой одинакова независимо от нуклеотидной последовательности, в действительности не совсем корректно. Детальный рентгеноструктурный анализ, построение моделей и термодинамические расчеты показали, что плоскости соседних пар оснований не строго параллельны. Каждая комплементарная пара оснований является как бы клином, отклоняющим ось спирали в одном или в другом направлении. Наибольший «крен» наблюдается тогда, когда два соседних аденина в одной цепи спарены с двумя тиминами другой. В этом месте происходит локальное искривление спирали. Если такие пары встречаются с периодичностью примерно один раз на 10 пар (т.е. один раз на каждый виток спирали), то молекула ДНК приобретает заметно искривленную форму. Изгибы в молекуле ДНК наблюдаются в тех участках последовательности, где с необычно высокой частотой встречаются повторы (А•Т)5–6, разделенные GC-богатыми участками из четырех-шести нуклеотидов. Биологическая роль искривления ДНК окончательно не установлена. Предрасположенность к такому изгибанию, зависящая от последовательности оснований, может иметь значение при наматывании молекулы ДНК на гистоновые октамеры в хроматине. Возможно, изгибание ДНК существенно и при специфическом связывании ДНК с белками в процессе регуляции экспрессии генов.
ДНК может находиться в линейной или кольцевой форме:
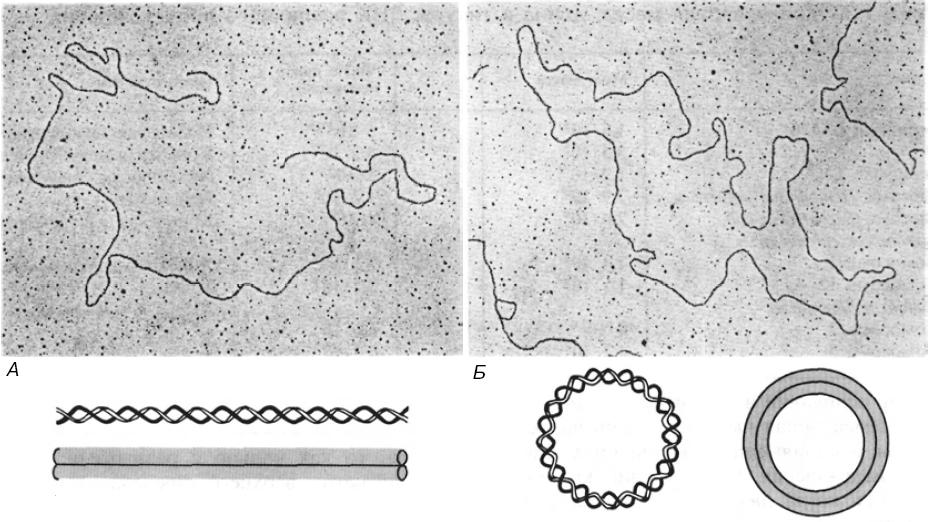 Схематическое
представление и электронные микрофотографии
линейной (А) и кольцевой (Б) двухцепочечной
ДНК фага λ
Схематическое
представление и электронные микрофотографии
линейной (А) и кольцевой (Б) двухцепочечной
ДНК фага λ
Бактериальные плазмиды, хромосомы некоторых бактерий, большинство митохондриальных и хлоропластных ДНК, геномы вирусов млекопитающих представлены единственной ковалентно замкнутой кольцевой дуплексной молекулой ДНК. Хромосома бактериофага λ на разных стадиях жизненного цикла существует то как линейная молекула, то как замкнутая кольцевая структура, то как кольцо с разрывами. По-видимому, никакого верхнего предела для размера кольцевой двухцепочечной молекулы ДНК не существует.
ДНК в клетке обычно находится в комплексе с белками. Связанный белок слегка раскручивает спираль ДНК, соответственно и число витков спирали на единицу длины становится меньше, чем у свободной В-ДНК. При удалении белка восстанавливается обычное число правозакрученных (положительных) витков спирали. В линейной молекуле ДНК это происходит достаточно легко, поскольку обе цепи свободно вращаются одна вокруг другой. В замкнутой же кольцевой молекуле общее число витков спирали топологически фиксировано, и число оборотов одной цепи вокруг другой не может быть изменено без компенсаторного образования витков противоположного знака где-нибудь в другом месте молекулы. Итак, когда естественные кольцевые дуплексы освобождаются от белков, с которыми они часто бывают связаны in vivo, происходит следующее:
1) число правозакрученных (положительных) витков спрали возрастает до величины, характерной для В-ДНК; 2) в самом дуплексе образуется столько же витков противоположного знака, чтобы компенсировать увеличение скрученности спирали.
О таких молекулах говорят, что они обладают отрицательной сверхспиральностью:
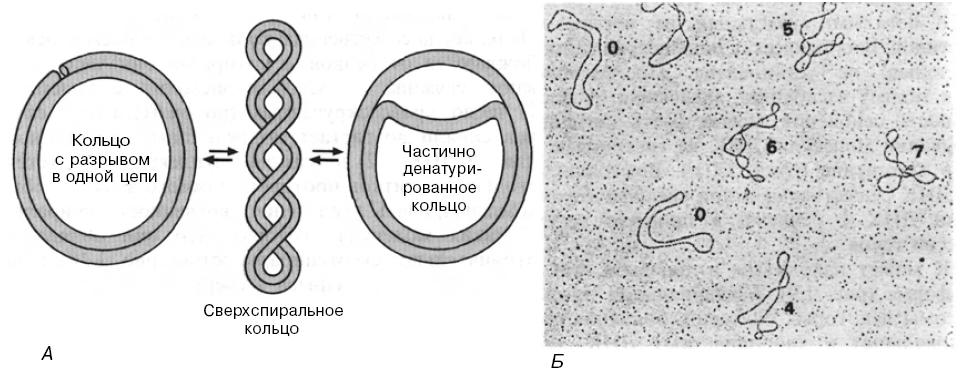
А. Схематическое изображение сверхспиральной кольцевой ДНК и релаксированных кольцевых форм, полученных либо в результате разрыва одной из двух цепей, либо в результате локального расхождения двух цепей Б. Двухцепочечная кольцевая ДНК фага М13 с разной степенью сверхспиральности. Цифрами обозначено число сверхвитков в каждой молекуле.
Упаковка ДНК в хромосомах
В клетках или вирусах ДНК, по-видимому, никогда не находится в свободной, вытянутой форме. Она связана с низкомолекулярными катионами — ионами двухвалентных металлов либо с ди- и полиаминами или белками, а возможно, с теми и с другими. Взаимодействие осуществляется с помощью электростатических сил — отрицательно заряженные фосфатные группы частично нейтрализуются положительно заряженными ионами металлов и полиаминами или основными аминокислотными остатками белков. В результате таких взаимодействий происходит конденсация ДНК с уменьшением объема, занимаемого молекулой, иногда в тысячу раз. Кольцевая ДНК Е. coli длиной 1,4 мм заключена в клетку, имеющую форму палочки диаметром 1 мкм и длиной 2 мкм; у эукариотических клеток ядерная ДНК длиной почти 2 м в стадии интерфазы заключена в ядре диаметром менее 10 мкм. Ядерная ДНК в клетках, находящихся в стадии митоза, конденсирована еще больше и в световом микроскопе имеет вид очень компактной структуры.
Упаковка ДНК в ядре
В средней эукариотической клетке общая протяженность геномной ДНК составляет около 2 м, диаметр ее ядра всего ~10-20 мкм. При этом совокупность генов, работающих в данной клетке, должна быть доступна для РНК-полимераз и транскрипционных факторов, а вся ДНК в делящихся клетках должна реплицироваться.
Сегодня известно, что упаковка ДНК в ядре эукариотической клетки осуществляется в несколько этапов . Сначала нить ДНК укладывается в нуклеосомы, при этом ее длина уменьшается в шесть-семь раз. Затем нуклеосомная нить складывается в так называемую 30 нм фибриллу (соленоид или зигзагообразную нить), что обеспечивает дополнительную компактизацию в 40 раз. Далее фибрилла организуется в большие (50 и более тысяч пар нуклеотидов) петли, концы которых закрепляются на белковом скелете ядра (его часто называют ядерным матриксом). На этом этапе линейные размеры ДНК сокращаются в 700 раз. Существуют и следующие уровни компактизации ДНК, информация о которых в настоящее время весьма скудна и противоречива.
Правильная упаковка ДНК с хромосомными белками осуществляется под наблюдением вспомогательных ферментов. Для корректировки упаковки ферменты-помощники используют энергию АТФ. Исследователи из Университета Пенсильвании (США) сумели искусственно воспроизвести сворачивание хромосомы, и, как говорят учёные, решающим фактором оказалась энергия — наличие в реакционной смеси молекул АТФ. Результаты экспериментов опубликованы в журнале Science.
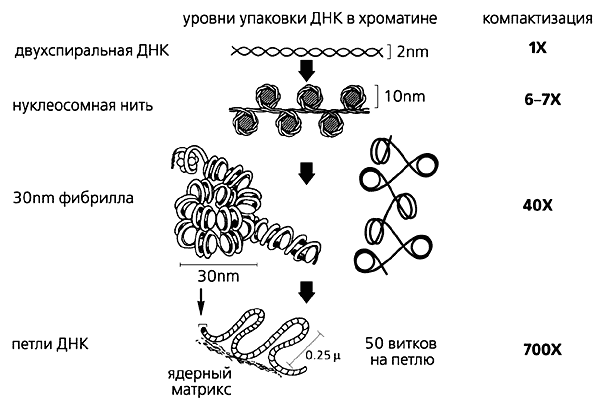
Рис. 1. Уровни упаковки ДНК в ядре эукариотической клетки.
Пока речь шла лишь об упаковке одной протяженной молекулы ДНК. В первом приближении таковой можно считать ДНК одной хромосомы. Однако геном эукариотической клетки разделен на несколько хромосом. Например, в клетках плодовой мушки дрозофилы - имеется четыре пары хромосом (в клетках человека их 46). Индивидуальные хромосомы можно увидеть под микроскопом только во время митоза. На остальных фазах клеточного цикла они не видны, и ядро клетки представляется относительно гомогенным. В течение многих лет молекулярных биологов интересовал вопрос, занимают ли отдельные хромосомы ограниченные пространства внутри ядра или же при декомпактизации хромосом ДНК каждой из них распределяется по всему ядру, неизбежно перемешиваясь с ДНК других хромосом. Около 10 лет назад ответ на этот вопрос был найден. Методы молекулярной гибридизации позволили окрашивать в интерфазном ядре индивидуальные хромосомы . Оказалось, что они, вопреки общепринятой в то время точке зрения, занимают внутри ядра ограниченные неперекрывающиеся пространства (названные "хромосомными территориями" и располагаются неслучайным образом: хромосомы, богатые генами, локализуются ближе к центру ядра, а бедные генами - ближе к его периферии. В поддержании специфических позиций хромосомных территорий важную роль играет ядерный матрикс.
Хромосомы эукариот
Хромосомы эукариотических клеток состоят в основном из хроматина — комплекса двухцепочечной ДНК и пяти гистоновых белков, обозначаемых H1, Н2А, Н2В, Н3 и Н4. Гистоны могут быть ацетилированы, метилированы, фосфорилированы, роlу(АDР)-рибозилированы, а гистоны Н2А и Н2В — ковалентно связаны с белком, называемым убиквитином. Какова роль воздействия указанных компонентов на структуру и функции гистонов — до конца не выяснено. Гистон H1 млекопитающих состоит из примерно 215 аминокислот; размеры других гистонов варьируют от 100 до 135 аминокислот. Все они содержат необычно большое количество положительно заряженной аминокислоты лизина; Н3 и Н4 отличаются от других тем, что у них достаточно высок уровень положительно заряженной аминокислоты аргинина. Соотношение между Н2А, Н2В, Н3 и Н4, содержащимися в хроматине низших эукариот (дрожжи, плесневые грибы), такое же, как в хроматине млекопитающих.
На электронно-микроскопических фотографиях в зависимости от условий выделения и степени растяжения хроматин выглядит либо как длинное волокно диаметром 10 нм, либо чаще как более вытянутое волокно с утолщениями — «бусинками» диаметром 10 нм, нанизанными по всей длине волокна с определенными интервалами:
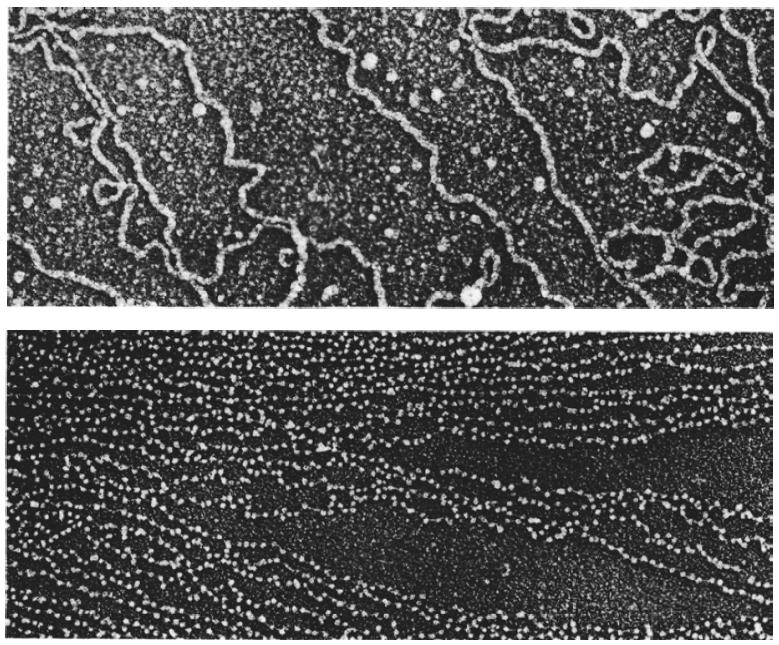
Электронные микрофотографии хроматина. А. Волокно хроматина диаметром 10 нм из почечных клеток CV1 обезьяны. Б. Хроматин из эритроцитов цыпленка, имеющий вид нити с нанизанными на нее бусинками.
Каждая из этих бусинок представляет собой нуклеосомный кор, на который намотан сегмент хромосомной ДНК длиной 145 пар оснований. Кор — это гистоновый октамер, состоящий из гистонов Н2А, Н2В, Н3 и Н4, по две молекулы каждого вида:
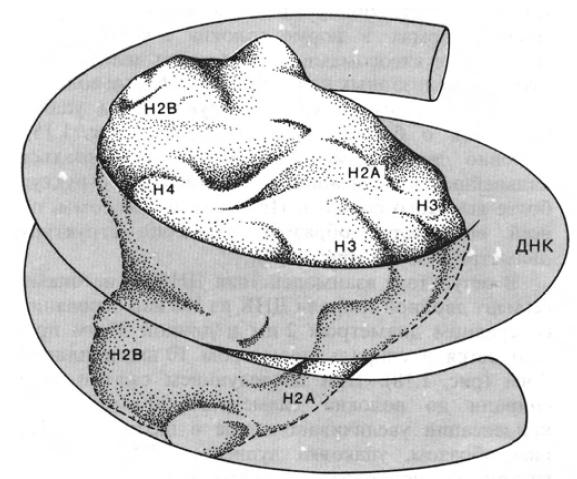
Модель нуклеосомного кора, построенная по данным кристаллографического анализа низкого и высокого разрешения. Сегмент ДНК (145 пар оснований), изображенный в виде трубки, обвивает гистоновый октамер, делая вокруг него 13/4 оборота
Молекула ДНК, обвиваясь 13/4 раза вокруг нуклеосомного кора, образует сверхспираль.
Пятый гистон, H1, не входит в состав нуклеосомного кора и не участвует в процессе наматывания ДНК на гистоновый октамер. Он контактирует с ДНК в тех местах, где двойная спираль входит и выходит из нуклеосомного кора:
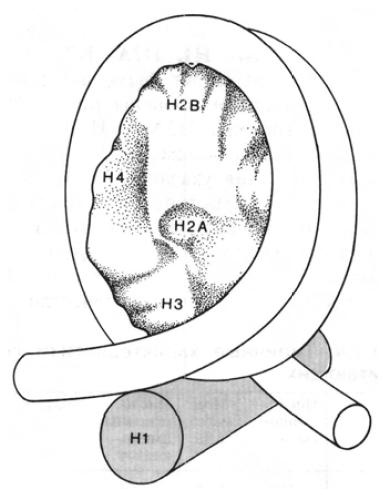
Гистон Н1 «сшивает» ДНК в местах, где она начинает и прекращает наматываться на нуклеосомный кор
В такой структуре с одним гистоновым октамером и молекулой гистона H1 ассоциированы 168 пар оснований спиральной ДНК. Как мы уже отмечали, на электронно-микроскопических фотографиях хроматин часто обнаруживается в двух альтернативных формах: в форме волокна с четко разделенными нуклеосомами (нуклеосомы имеют вид бусинок, нанизанных на нитку) или в форме волокна диаметром 10 нм, в котором нуклеосомы упакованы бок о бок по всей его длине. Волокно диаметром 10 нм может подвергаться дальнейшей конденсации с образованием структур более высокого порядка. При этом нуклеосомы, по всей видимости, образуют соленоид — структуру диаметром 30 нм:
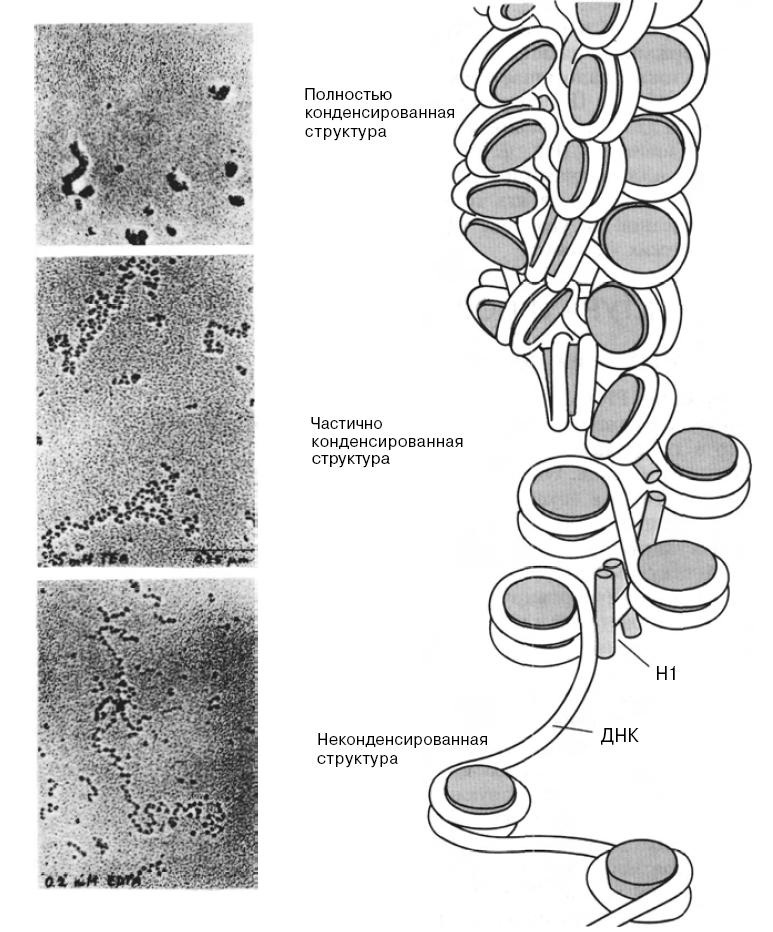 Структура
хроматина с разной степенью конденсации.
В
нижней части рисунка представлен
хроматин, находящийся в растянутой
форме; он имеет вид нити с нанизанными
на нее бусинками.
Далее изображен
хроматин в частично конденсированной
форме, представляющий собой волокно
диаметром 10 нм.
В верхней части
рисунка представлен хроматин в наиболее
конденсированном состоянии, когда
волокно диаметром 10 нм образует соленоид
диаметром 30 нм.
Обратите внимание
на взаимодействие молекул гистона Н1,
связанных с каждой нуклеосомой, которое
способствует конденсации волокна
диаметром 10 нм в более плотную структуру
Структура
хроматина с разной степенью конденсации.
В
нижней части рисунка представлен
хроматин, находящийся в растянутой
форме; он имеет вид нити с нанизанными
на нее бусинками.
Далее изображен
хроматин в частично конденсированной
форме, представляющий собой волокно
диаметром 10 нм.
В верхней части
рисунка представлен хроматин в наиболее
конденсированном состоянии, когда
волокно диаметром 10 нм образует соленоид
диаметром 30 нм.
Обратите внимание
на взаимодействие молекул гистона Н1,
связанных с каждой нуклеосомой, которое
способствует конденсации волокна
диаметром 10 нм в более плотную структуру
В результате взаимодействия ДНК с гистонами сегмент двойной спирали ДНК из 168 пар оснований со средним диаметром 2 нм и длиной 57 нм превращается в спираль диаметром 10 нм и длиной 5 нм. При последующем сжатии этой спирали до волокна диаметром 30 нм степень конденсации увеличивается еще в шесть раз. Таким образом, упаковка дуплекса ДНК с пятью гистонами приводит к 50-кратной конденсации ДНК. Однако даже столь высокая степень конденсации не может объяснить почти 5000-кратное уплотнение ДНК в метафазной хромосоме.
Эукариотический хроматин содержит и другие белки, которые обычно называют негистоновыми. Некоторые из них, например ферменты, необходимые для репликации и экспрессии ДНК, могут связываться с хроматином временно. Белки, принимающие участие в различных процессах регуляции, связываются с ДНК только в специфических тканях или на определенных стадиях дифференциации.
Сегодня пришли новые технологии и методы, благодаря чему микроскопия в биологии стала трехмерной. Появилась возможность рассмотреть хромосому в интерфазном ядре и получить информацию о локализации в нем сразу всех хромосом человека . Для этого широко применяют гибридизацию in situ (FISH) ДНК индивидуальных хромосом, меченной флуоресцентными красителями, с ДНК интерфазного ядра. Затем с помощью лазерного сканирующего микроскопа получают серию оптических срезов ядра, на которых зарегистрированы интересующие исследователя сигналы. Такие оптические срезы можно рассматривать отдельно, использовать для создания ортогональных проекций или для реконструкции трехмерной организации клеточного ядра.
Хромосомы прокариот
Насколько известно, в упаковке прокариотической геномной ДНК участвуют только два или три белка. О природе взаимодействия этих белков с ДНК и о структуре конденсированного комплекса белокнуклеиновая кислота известно немного. У Е. coli, по-видимому, существует лишь один белок или один класс ДНК-связывающих белков, называемых HU-белками; по своему размеру, содержанию лизина и аргинина, антигенным свойствам они сходны с эукариотическим гистоном Н2А. Другой белок, белок II, обнаруженный у Е. coli и цианобактерий, по повышенному содержанию лизина и ДНК-связывающим свойствам также напоминает эукариотический гистон. Белки HU и II обнаружены в количествах, достаточных для образования комплекса по крайней мере с половиной ДНК Е. coli и, по-видимому, совместно с полиаминами и еще неизвестными нам белками могут осуществлять те же самые функции при конденсации и упаковке ДНК, что и пять эукариотических гистонов.
Митоз
Митоз, или непрямое деление, - основной способ размножения эукариотических клеток, обусловливающий, в частности, возможность увеличения их биомассы, рост и регенерацию. Митоз состоит из четырех фаз.
Первая – профаза – характеризуется началом цикла компактизации хромосом, который продолжается в течение всей этой фазы. Вследствие этого хромосомы становятся видимыми под микроскопом, причем уже в средней профазе митоза они представляются двойными структурами – сестринскими хроматидами, которые являются таковыми, пока удерживаются центромерой вместе. К концу профазы исчезают ядрышко и ядерная мембрана.
Вторая –метафаза. Процесс компактизации хромосом продолжается и ведет к еще большему укорочению их длины. Хромосомы выстраиваются по экватору клетки. Хроматиды соединены между собой между собой в центромере, называемой также первичной перетяжкой. Появляются нити митотического веретена, которые присоединяются к ценромерам. Каждая ценромера испытывает напряжение, поскольку нити веретена тянут ее к противоположным полюсам.
Полюса клетки формируются специальными органеллами – центросомами.
Третья – анафаза – начинается с разрыва ценромеры, в результате чего сестринские хроматиды расходятся к разным полюсам клетки. С этого момента каждая пара сестринских хроматид получает название дочерних хромосом.
Четвертая – телофаза. Хромосомы достигают полюсов клетки, появляются ядерная мембрана, ядрышко. Происходят декомпактизация хромосом и восстановление структуры интерфазного ядра. Заканчивается митоз делением цитоплазмы и в типичных случаях – восстановлением исходной биомассы дочерних клеток.
Биологическая роль митоза состоит в обеспечении идентичной генетической информацией двух дочерних клеток. Это достижимо только благодаря циклу компактизации – декомпактизации, который и позволяет распределить наследственные молекулы в минимальном объеме митотических хромосом. В противном случае, учитывая размеры клетки (десятки или сотни кубических микрометров) и длину декомпактизованной хромосомы (сантиметры), каждое клеточное деление сопровождалось бы хаотичным переплетением хромосомного материала.
В эволюции эукариотических клеток, видимо, это обстоятельство и послужило причиной становления столь сложного генетического процесса, как митоз.
Каждая хромосома индивидуальна, т.е. характеризуется свойственными только ей размерами, формой и положением центромеры. В клетках тела организмов, размножающихся половым путём, любая хромосома представлена двумя копиями, или гомологами . При образовании половых клеток в мейозе в каждую из них попадает одна из двух гомологичных хромосом . При оплодотворении парность гомологичных хромосом восстанавливается: одна хромосома каждой пары отцовская, другая – материнская. Совокупность признаков хромосомного набора (число хромосом, их размер и форма) постоянна для клеток каждого вида и называется его кариотипом. В кариотипе различают пару определяющих пол организма половых хромосом и все остальные хромосомы – аутосомы. Изучением поведения хромосом в митозе и мейозе, а также роли хромосом, особенно половых, при передаче признаков от одного поколения к другому привело к созданию в нач. 20 в. хромосомной теории наследственности и до настоящего времени исследуется огромным количеством как цитогенетиков, так и других ученых, включая и физиков. Как уже было сказано, хромосомой часто называют генетический материал бактерий и вирусов, хотя его строение отличается от хромосом эукариотических организмов.