

Цепной условный рефлекс
Рассмотрим, как формируется условный рефлекс на цепь раздражителей (рис. 4.2). В первое время образования условного рефлекса на цепь раздражителей устанавливаются частные условные связи (а, б, в) между его компонентами (А, Б, В) и пищевым (П) «центром». После более или менее длительной тренировки рефлекса эти связи затормаживаются и образуется общая связь (г) между возникшим в результате синтеза отдельных раздражений функциональным комбинационным центром (Г) и корковым представительством пищевого (П) «центра» (см. рис. 4.1, II). Эта схема, предложенная Л. Г. Ворониным, составлена на основе опытов, по результатам которых был сделан вывод, что вначале каждый из последовательно действующих раздражителей {А, Б, В) устанавливает связь с корковым представительством пищевого «центра» (П), благодаря образованию параллельных {а, б, в) связей. Затем, после некоторого периода тренировки рефлекса, компоненты цеп-
92
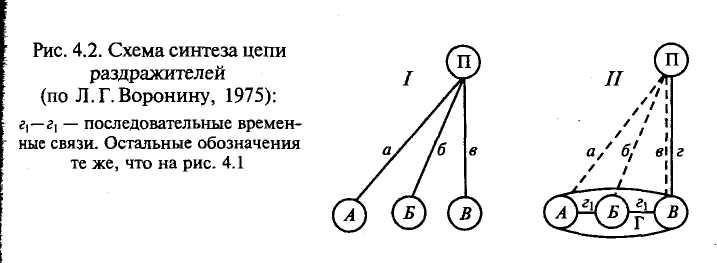
ного раздражителя синтезируются в целостную систему (Г) с определенной последовательностью их действия. Параллельные связи при этом затормаживаются, а вместо них образуется последовательная связь (zi — гО комплекса раздражений с пищевой деятельностью (см. рис. 4.2, II). Однако параллельные связи не исчезают совсем, а только затормаживаются. Например, они растормаживаются при повышении пищевой или общей возбудимости (голодание, действие фармакологических веществ). В таких случаях компоненты сложного раздражителя в течение некоторого времени снова будут вызывать условный рефлекс.
Возможность получения условного рефлекса на цепь раздражителей зависит от филогенетического уровня развития нервной системы данного вида животного. Так, у обезьян (макак, павианов, капуцинов), как показали опыты Л. Г. Воронина (1958), после 40— 200 применений цепного раздражителя его компоненты, испытанные в отдельности, в большинстве случаев не вызывают условного рефлекса. У низших позвоночных (рыбы, рептилии) даже после 700—1300 применений цепи раздражителей ее компоненты сохраняют сигнальное значение. У этих животных условный рефлекс на цепь раздражителей вырабатывается довольно легко, но сложный раздражитель не становится единым: каждый из его компонентов сохраняет свое сигнальное значение. На основании данных опытов Л.Г.Воронин пришел к заключению, что у высших животных в результате упрочения условного рефлекса на сложные раздражители создается функциональный комбинационный центр (ФКЦ). Подобное мнение высказывал еще И.П.Павлов, который отмечал, что между ФКЦ и безусловным «центром» устанавливается временная связь. Вопрос о местоположении этого «центра» относится к проблеме локализации временной связи, и к этому вопросу мы еще вернемся.
Цепные двигательные условные рефлексы
У высших млекопитающих (кошек, собак, обезьян и др.) можно довольно легко вырабатывать однородные (с одного эффектора) или разнородные (с нескольких эффекторов) цепные двига-
93
тельные условные рефлексы. Этот процесс схематически показан на рис. 4.3. На схеме показано, что на раздражитель А выработано 7-е, на раздражитель Б — 2-е и на раздражитель В — 3-е условно-рефлекторное движение. Затем если раздражители свести в цепь А— Б—В, то в соответствии с их последовательностью осуществятся условные движения 1, 2, 3. Как показали исследования, в начале выработки главную роль играют прямые условные связи между внешним раздражителем и выработанным на него движением (А-> 1, 7э—»2, В-+3). В дальнейшем, по мере синтеза раздражителей и движений, ведущее значение приобретают последовательные связи: Л—>7—»2—¥ 3—>П. Поэтому, как показали специальные исследования, после достаточно длительной тренировки достаточно применить первый компонент цепи раздражителей (А), чтобы осуществилась вся цепь движений (7—»2-) 3). В каких отношениях находятся внешняя и кинестезическая сигнализации в случае формирования цепи движений на цепь раздражителей? Можно предположить (рис. 4.4), что если внешний раздражитель (А) вызывает движение (7), то в двигательно-кинестезическую область коры поступают импульсы, создающие там очаг возбуждения (а). При постоянном сочетании очагов возбуждения, возникающих от экстероцептив-ного раздражителя (Л) и от проприоцептивного (а), создается условный комбинационный центр (Аа). Таким же образом возникает центр Бб вследствие сочетания раздражителя Б и движения (2). Когда такие синтезированные очаги возбуждения (Аа, Бб, Вв и т.д.), вызывающие соответствующие им движения (7, 2, 3), сочетаются многократно и в определенной последовательности, то в конечном счете это внешне выглядит так, что цепь экстероцеп-тивных раздражителей (А, Б, В) всегда вызывает цепь движений (7, 2, 3). На самом же деле, как это следует из схемы, цепь движений осуществляется в ответ на цепь комбинаций экстероцептив-ных и проприоцептивных раздражителей (Аа, Бб, Вв). При этом ведущее значение приобретают либо экстероцептивные раздражения, либо первое звено этих движений и цепь кинестезических
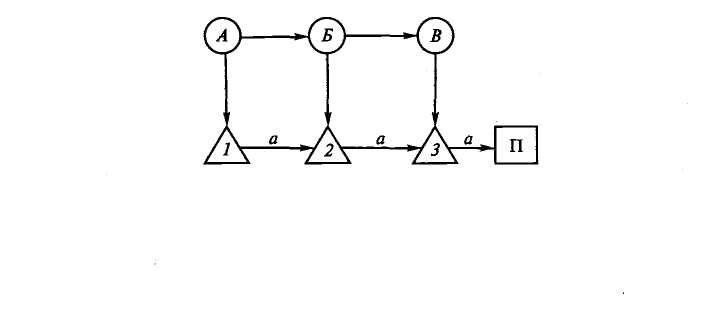
путем объединения отдельных двигательных условных рефлексов
(по Л. Г. Воронину, 1975):
А, Б, В — экстероцептивные условные раздражения; 1, 2, 3 — двигательные акты; П — подкрепление; а — временные связи
94
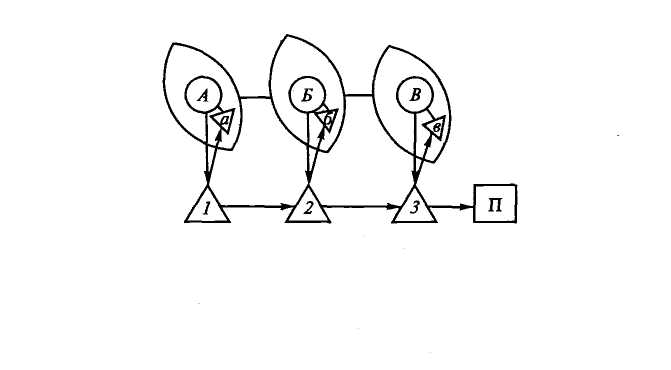
центров» в случае цепи условно-рефлекторных движений
(по Л.Г.Воронину, 1975):
а, б, в — кинестезические раздражения. Остальные обозначения те же,
что на рис. 4.3
раздражений (А—>а—> £—»в). В первом случае это будет цепь движений 1, 2, 3, соответствующая своей последовательностью цепи условных раздражителей АБВ; во втором случае может быть выработано три варианта условно-рефлекторных ответов: на раздражитель А — цепь движений 1—> 2-> 3, на раздражитель Б — цепь движений 2—>3, на раздражитель В — движение 3. Это значит, что одна система рефлексов, вызываемая в определенной последовательности системой раздражителей, может быть разбита на три самостоятельных условных рефлекса благодаря тому, что производилось подкрепление реакций, появляющихся раньше «опережающих» их раздражителей.
У человека также проявляется тенденция к синтезу отдельных движений, сведенных в цепь. Это выражается в том, что испытуемый может совершить движение раньше действия сигнала движения. Однако тенденция опережения подавляется либо инструкцией, которую давали испытуемому, — производить движения в соответствии с их сигналами, либо его собственным опытом того же содержания. Такого рода синтез движений в единую цепь, осуществляющийся только на первый «пусковой» компонент цепи раздражителей, возможен при специальной инструкции или на основании личного опыта испытуемого, в результате чего он знает о возможности совершать движение, не дожидаясь сигнала.
Формирование изображенной на схеме системы двигательных рефлексов у животных достигается только в том случае, когда последовательно возникающие, например, три движения (1—>2—> 3) в ответ на первый компонент (А) цепи раздражителей подкрепляются пищей. Если же подкреплять цепь движений только тогда, когда каждый из ее компонентов соответствует условному эксте-роцептивному раздражению, то животное не станет производить движения, пока не подействует этот раздражитель. Так, например, если после осуществления движения 1 (см. рис. 4.4) в ответ на
95
раздражитель А не включать раздражители Б, В, то животное не! будет производить движения 2, а начнет в силу иррадиации пище-] вого возбуждения беспокойно двигаться и совершать пищевые по-] иски. Связь между отдельными движениями и раздражителями на-1 глядно обнаружилась в опытах Г. И. Ширковой на шимпанзе (ла-1 боратория, руководимая Л. Г. Ворониным). Так, обезьяна в подоб- { ном случае не производила движения 2, а рукой стучала по окош- \ ку (рис. 4.5), где обычно появлялся свет (раздражитель Б), как бьн «требуя» включения очередного раздражения. Из этого эксперимента был сделан вывод, что при образовании таким способом цепного двигательного рефлекса каждый из компонентов цепного ~> внешнего раздражителя, за исключением первого, приобретает двойное значение: он становится сигналом последующего движения и подкреплением уже совершенного движения.
Известно четыре способа образования цепных условных рефлексов у животных. Выше был описан первый способ — объединение в цепь экстероцептивньгх одиночных раздражителей одиночных двигательных реакций (Л.Г.Воронин, 1947). Второй способ, предложенный А. В. Напалковым (1956), заключается в наращивании цепи движений с подкрепляемого конца. Например, сначала обучают животное (голубя, крысу и т.д.) по условному сигналу (включение лампочки) клевать (нажимать) первую полку в экспериментальной камере. Затем, пустив достаточно голодное животное в камеру, не подают условного сигнала, заставляя животное совершать поисковые реакции. На вторую полку кладут приманку. Как только животное прикоснется ко второй полке, сразу же включают лампу (условный сигнал), и после клевания (нажимания) второй полки животное получает пищевое подкрепление.

В результате нескольких таких сочетаний у животного закрепляется клевание (нажимание) второй полки. После этого вводят Другой эк-стероцептивный сигнал — включение звонка, предшествующее клеванию (нажиманию) второй полки. Таким образом образуется двучленная, трехчленная и т.д. цепь движений. В противоположность указанному способу при третьем способе формирования цепи двигательных рефлексов осуществляется «вклинивание» новых движе-
Рис. 4.5. Шимпанзе стучит ний и раздражителей подобным же по окошку, «требует» вклю- образом, но между последним зве- чения сигнала ном цепи и подкреплением. Нако-
96
со Л. Г. Воронину, 1965) нец, при четвертом способе фор-
тонирования цепи движений животное не ограничивают в движениях, но подкрепляют только те цепи, которые являлись «правильными». Оказалось, что в таких условиях, например, обезьяны достаточно быстро научались производить нужную цепь движений, а се ненужные действия у них постепенно исчезали. Синтезирова--ние цепи движений происходило с подкрепляемого конца, так же как и при втором способе.
У животных цепи движений вырабатываются с различной степенью трудности в зависимости от филогенетического уровня развития нервной системы. У черепах, например, в течение длительного ^времени с большим трудом можно выработать весьма нестойкую трехчленную цепь движений, у голубей удается образовать довольно прочную цепь из 8 — 9 движений, а у млекопитающих — из еще большего числа движений. Был сделан вывод, что существует зависимость скорости образования отдельных звеньев и всей цепи движений в целом от уровня филогенеза животного. Как видно из данных табл. 4.1, у черепах для образования каждого нового компонента цепи движений требуется больше сочетаний, чем для выработки предыдущего компонента. У голубей это явление выражено в меньшей степени, у крыс обнаруживается слабая тенденция к такой закономерности, а у кошек и собак она полностью отсутствует. По мнению Л. Г. Воронина, этот факт говорит о том, что у животного с более развитой филогенетически нервной системой тренировка временных связей играет большую роль. По-видимому, она лежит в основе так называемого переноса опыта на другую или аналогичную ситуацию.
Таблица 4.1
Образование отдельных звеньев и цепи условных движений
(по Л.Г.Воронину, 1965)
Объект |
Число сочетаний, потребовавшееся для образования условных рефлексов (средние цифры) |
||||
исследования |
1 -членного |
2-членного |
3-членного |
4-членного |
5-членного |
Черепаха |
8 |
58 |
116 |
Не вырабатывается |
|
Голубь |
10 |
16 |
15 |
24 |
30 |
Кролик |
15 |
18 |
14 |
18 |
21 |
Крыса |
9 |
12 |
13 |
28 |
18 |
Кошка |
21 |
15 |
28 |
24 |
18 |
Собака |
11 |
12 |
15 |
12 |
14 |
Человек |
2 |
1 |
1 |
1 |
1 |
4 Шул
97
С Исключительной легкостью образуются цепи движений у человека. Испытуемый после первого опыта обычно говорит, что он сразу догадался, какое движение нужно производить, чтобы решить задачу. Выработанную таким образом у человека или животного цепь движений можно изобразить как Ах Б2 Въ ГА -> П (буквами обозначаются в этом случае сигналы, а цифрами — движения). Цепь движения в опытах с животными подкрепляется (П) пищей, а в опытах с человеком — сигналом, заранее обусловленным инструкцией, например звуком определенной частоты.
Автоматизация условных рефлексов
Очень многие условные рефлексы у животных и человека после длительной тренировки автоматизируются, становятся как бы независимыми от остальных проявлений высшей нервной деятельности. Автоматизация, как правило, развивается постепенно. Вначале она может выражаться в том, что отдельные движения опережают соответствующие сигналы. Затем наступает период, когда цепь движений полностью осуществляется в ответ на первый, «пусковой», компонент цепи раздражителей. При первом взгляде на результат тренировки условного рефлекса может создаться такое впечатление, что сначала рефлекс «привязан» к чему-то, управляющему им, а затем после длительного упражнения он становится до некоторой степени независимым. Именно до некоторой степени, потому что как только нарушится обстановка, при которой условный рефлекс образовался и в дальнейшем функционирует, это «что-то» снова «направляет» условный рефлекс в определенное русло. Например, у собаки на цепь сигналов выработана цепь движений. После длительной тренировки животное совершает эту цепь в ответ только на первый компонент цепи раздражений, не ожидая действия всей цепи. Если же вдруг происходит изменение в цепи сигналов, то совершившаяся теперь цепь движений не приводит к достижению пищи. Что же произошло? С физиологической точки зрения, произошло нарушение установившегося ранее взаимоотношения, «согласованности» сигналов и деятельности организма, стереотипии. Как правило, изменение в цепи сигналов вызывает ориентировочный рефлекс. В конечном счете это способствует установлению новых отношений между сигналами и ответом организма на них.
Наступление баланса системы условного—безусловного возбуждения (или торможения) при воздействии на нервную систему внешней и внутренней среды организма И.П.Павлов оценивал как «системность в работе головного мозга». Взгляд на саморегуляцию функции организма как на процесс сопоставления нервных состояний и обстоятельств, вызывавших их, отражает реальные явления, но далеко не во всех случаях здесь можно назвать конкрет-
98
it механизмы. Из всего сказанного следует, что воздействие внеш-ей и внутренней среды на организм создает в головном мозге со-ояние, которое можно описать системой очагов возбуждений и орможений.
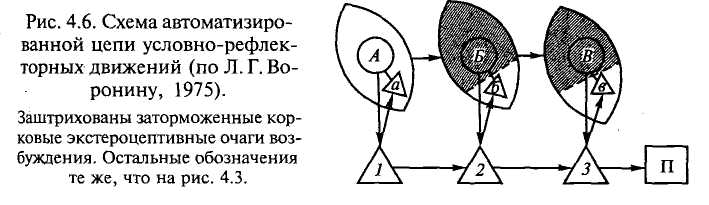
Как же можно представить физиологические механизмы хоро-) известного явления автоматизации двигательных условных реф-ексов? Если на организм животного действует система последо-ательно появляющихся внешних раздражений, то в соответствии этим в мозгу возникает система очагов возбуждения, причем 1ый объединяется с очагом возбуждения от кинестезических цражений (см. рис. 4.4). Объединенный очаг возбуждения связы-ается с очагом пищевого возбуждения. Если таких комплексных очагов несколько и они возникают последовательно друг за другом, то создаются условия для цепной реакции. Поскольку каж->ш из комплексных очагов возбуждения вызывает движение, все очаги, возникающие друг за другом, вызывают цепь движений, приводящих в конечном счете к появлению пищи или к устранению помех на пути к пище. В результате длительной тренировки цепи движений наступает ее автоматизация, выражающаяся в том, что вся цепь осуществляется в ответ только на первый компонент цепи раздражителей. Это происходит вследствие того, что возбуждение, сконцентрированное в районе данной временной связи, получило там возможность быстро распространяться по уже готовым связям при незначительном «толчке» извне — первом сигнале в цепи. Очевидно, между очагами возбуждения и торможения в силу постоянства условий тренировки временных связей устанавливается такое отношение, которое может «освободить» какую-либо систему временных связей от влияния других систем. В этом случае очаги кинестезического возбуждения при помощи индукционного торможения «разобщаются» с очагами возбуждения других афферентаций. На рис. 4.6 изображена приведенная ранее схема комплексных «центров» цепного двигательного рефлекса. На ней показано торможение (заштриховано) всех экстероцептивных очагов возбуждения, за исключением первого «пускового», которое может быть легко устранено (растормаживание), например, возникновением ориентировочного рефлекса при изменении установившихся отношений между раздражителем и организмом.
99
АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА
Для понимания механизмов аналитико-синтетической деятельности мозга имеют большое значение представления И. П. Павлова о коре как о мозаике из тормозных и возбужденных пунктов и его понимание функции коры как динамической системы этих пунктов. П. С. Купалов, ученик И. П. Павлова, при изучении условных рефлексов с кожного анализатора обнаружил, что «очаги» возбуждения и торможения в коре могут быть строго разграничены. Он выработал чередующиеся положительные и отрицательные условные рефлексы на ряд касалок, приклеенных вдоль туловища собаки. При этом каждая нечетная касалка была положительной, а каждая четная — отрицательной. После того как касалки (начиная со второй и кончая восьмой) приобрели свое сигнальное значение, были испытаны остававшиеся до сих пор без применения первая и девятая касалки. Оказалось, что они вызвали положительный эффект с первого же применения, «с места», хотя находились в районе действия второй и восьмой касалок, т. е. отрицательных раздражений.
Дальнейшие исследования выявили другой интересный факт: каждая из впервые примененных касалок вызывала эффект в зависимости от того, была ли она четная или нечетная. Так, например, если были выработаны условные рефлексы на восемь касалок (каждая нечетная — положительная, а четная — отрицательная), то впервые испытанные девятая и одиннадцатая касалки вызывали условное слюноотделение, а десятая и двенадцатая оставались без ответа. На основании рассмотренных опытов И. П. Павлов пришел к выводу, что «кора полушарий должна представлять собой грандиозную мозаику, грандиозную сигнализационную доску. На этой доске постоянно остаются огромные запасы пунктов для образования новых сигнальных условных раздражителей, а кроме того, занятые ранее пункты более или менее часто подвергаются изменениям в их связях с разными деятельностями организма, в их физиологическом значении»1.
Вслед за открытием корковой мозаичности, образующейся довольно легко, в павловских лабораториях стали исследовать явления корковой системности. Этот феномен проявлялся в виде процесса объединения «пунктов» возбуждения и торможения, что представляло уже, говоря словами И.П.Павлова, «значительный нервный труд для животных». Так, в специальных экспериментах обнаружилось, что если у собаки долгое время применять одну и ту же систему рефлексов, а затем изменить в ней порядок следования раздражителей, то у некоторых животных возникает невротическое состояние, выражающееся в резком уменьшении, а затем
1 Павлов
И.П. Лекции
о работе больших полушарий головного
мозга. Поли. собр. соч.
- М.; Л.: Изд-во АН СССР, 1951. - Т. IV.
— С. 231.
Павлов
И.П. Лекции
о работе больших полушарий головного
мозга. Поли. собр. соч.
- М.; Л.: Изд-во АН СССР, 1951. - Т. IV.
— С. 231.
100
исчезновении всех условных рефлексов. Вместе с тем если сохраняется постоянная система раздражителей, то животное способно решить очень трудную задачу.
Следующий пример иллюстрирует это положение. У собаки была выработана система из 6 слюнных условных рефлексов, следующих друг за другом через равные шестиминутные интервалы. Один из рефлексов был отрицательным, а остальные — положительными. После того как рефлексы были упрочены, в систему из раздражителей был введен новый агент, который применялся в течение опыта четыре раза. Он действовал каждый раз через 6 мин после какого-либо из условных раздражителей системы, но подкреплялся пищей только при четвертом применении. Выработка системы условных рефлексов протекала с большим трудом, что выразилось в сильном возбуждении животного и его отрицательном отношении к экспериментальной обстановке. В конечном счете эта система рефлексов все же была выработана: каждый из старых раздражителей по-прежнему вызывал условный рефлекс, а на новый агент следовал ответ только при его четвертом применении.
Таким образом, сигнальным признаком стало место этого агента в системе раздражителей, порядок его следования. Когда система стала прочной, было произведено превращение, или, как говорят в лаборатории, «переделка» в новый стереотип. Теперь, наоборот, первые три применения раздражителя сопровождали пищей, а четвертое нет. Животное снова бурно реагировало на необычное соотношение положительных и отрицательных раздражителей, но затем изменило свои реакции в соответствии с новым значением сигналов. В связи с тем, что система, или стереотип, условных рефлексов может меняться в соответствии с системой раздражителей, само явление было названо динамической стереотипией.
Исследованием Э.А.Асратяна (ученик И.П.Павлова) было обнаружено, что лучшим способом выявления системности служит замена всего стереотипа раздражителей одним из агентов, входящих в него. Обычно при образовании динамического стереотипа чем сильнее раздражитель, тем больше условное слюноотделение (в соответствии с законом силы). Оказалось, что если стереотип раздражителей заменить одним из его компонентов — светом, то этот слабый раздражитель вызывает не соответствующую ему величину условного рефлекса, а ту, которую вызывал замененный им раздражитель. Например, на месте дифференцировочного раздражителя другой сигнал останется либо без эффекта, либо слюноотделение будет незначительным. Стереотип рефлексов воспроизводится более отчетливо при замене системы сигналов слабым раздражителем. Объясняется это тем, что сильный агент не дает проявиться следам раздражений прежнего стереотипа. Кроме того, Э.А.Асратян обнаружил, что применение условного раздражителя, не входящего в данный стереотип, но выработанного на базе
101
того же безусловного рефлекса, также позволяет выявить системность в работе головного мозга. Интересно, что стереотипность может проявляться и по электроэнцефалографической характеристике функционального состояния мозга. Так, в лаборатории Л. Г. Воронина у кроликов, находящихся в экспериментальной камере, наблюдали появление, казалось бы, без всяких на то причин групп синхронизированных колебаний электрической активности мозга (рис. 4.1>- Они стали возникать после того, как животное несколько раз участвовало в опытах с «навязыванием ритма» (стимуляция мелькающим светом). Очевидно, обстановка опыта, представляющая собой стереотип, систему раздражителей, неоднократно совпадавшая с мелькающим светом, ритм которого «навязывался» электрической активности головного мозга кролика, в дальнейшем стала условным сигналом этой реакции и вызывала ее даже в отсутствие специальных раздражений.
Несмотря на динамичность деятельности головного мозга, проч-
ная система временных связей может сохраняться более или менее длительно, препятствуя образованию нового стереотипа реакции. Так, П.С.Купалов (ученик И.П.Павлова) обнаружил, что после выработки у собаки прочной системы рефлексов на чередующиеся положительные и тормозные раздражения касалками долгое время не удается выработать условный рефлекс на светящийся круг. После образования рефлекса круг воспроизводил старый стерео-
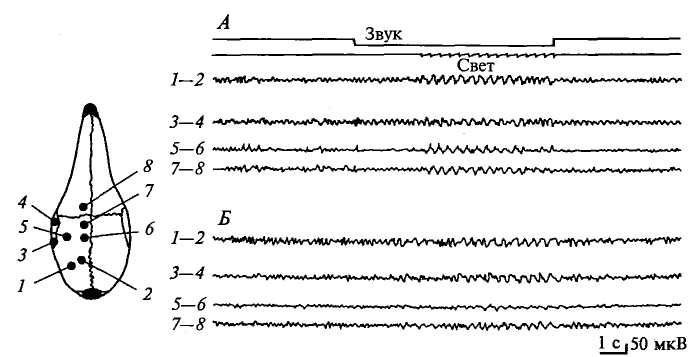
Рис. 4.7. Межсигнальная синхронизация электрической активности
мозга кролика:
А — световой раздражитель, вызывающий наведенный ритм в ЭЭГ; Б — межсигнальная синхронизация в ЭЭГ; 7—2 — ЭЭГ зрительной области коры; 3—4 — ЭЭГ слуховой области коры; 5—6 — ЭЭГ кожно-кинестезической области коры; 7— 8 — ЭЭГ двигательной области коры. Положение эпидуральных электродов показано на схеме
102
тип, поочередно вызывая слюноотделение то меньшей, то большей величины.
Старый стереотип может восстановиться и тогда, когда после образования нового стереотипа в экспериментах сделан, например, 10-дневный перерыв. В павловских лабораториях было обнаружено, что после перерыва замена системы раздражителей одним агентом вызывала не адекватный ему эффект, как это было до перерыва, а в зависимости от места его применения в опыте. Другими словами, здесь воспроизводился старый стереотип условных рефлексов. Интересно, что растормозить старый стереотип может не только сильное возбуждение, но он восстанавливается и при развитии, например, сонного торможения. Из результатов приведенных экспериментов был сделан вывод, что реальным условным сигналом служит комбинация из внешнего агента и различных внутренних состояний нервной системы, приуроченных к определенному отрезку времени. Этим объясняется, почему условные рефлексы при стереотипном применении раздражителей образуются несколько легче, чем в том случае, когда промежутки времени между ними непостоянны.
Э.А.Асратян показал, что при стереотипном применении раздражителей положительный раздражитель, действующий перед дифференцировочным агентом, вызывает уменьшенный эффект. Этот феномен Асратян назвал «ретроградным» (обратным) торможением и объяснил его взаимоиндукционными отношениями стойких очагов возбуждения и торможения, возникающих при образовании стереотипа. Согласно его точке зрения, такие отношения проявляются даже в том случае, если действует только положительный раздражитель (в силу стереотипии). Явление «ретроградного» торможения наблюдали Л. Г. Воронин, Л. Н. Норкина и другие ученые при исследовании стереотипа двигательных условных рефлексов у обезьян: величина двигательного ответа на положительный условный раздражитель, примененный перед дифференцировочным, была несколько уменьшена, а латентный период увеличен. Часто раздражитель оставался без ответа. «Ретроградное» торможение условного рефлекса было быстропроходя-щим. Оно наблюдалось только в период становления стереотипа, затем проявлялось все реже и, наконец, совсем исчезало. Поэтому его трудно было связать с наличием стойких очагов возбуждения и торможения стереотипа. Л.Г.Воронин объясняет «ретроградное» торможение как следствие «генерализованного» условного рефлекса на интервал времени между раздражителями стереотипа и, в частности, времени действия дифференцировочного раздражителя. По-видимому, сначала, пока еще не наступила специализация условного рефлекса на время, торможение (действие тормозного сигнала) возникает в широком временном диапазоне. После специализации условный рефлекс, на время «сливаясь» с рефлексом на
103
специальный раздражитель, становится незаметным, «ретроградное» торможение исчезает. Это предположение находит подтверждение в характеристике условного рефлекса на время действия стереотипных раздражителей. По данным Э. А. Асратяна, по мере тренировки стереотипа условный рефлекс на время перестает проявляться, а главное значение приобретают наличные раздражители и порядок их объединения в систему.
Физиологические механизмы многих явлений обыденной жизни человека, которые называются привычками, навыками, могут быть поняты в свете экспериментальных данных павловской школы о динамическом стереотипе. Любой укрепившийся навык у человека в своей основе имеет систему временных связей, которая нарушается только после изменения условий ее образования. Каждый из нас может привести множество таких примеров из своей жизни. Например, если в комнате перенесли выключатель электросети с правой стороны от двери на левую, то в первый день человек протягивает руку к тому месту, где раньше был выключатель, но потом вспоминает, что перемена произошла еще вчера по его же просьбе. Прочность навыка и индивидуальные особенности человека определят, как долго удержится привычка. Возможно, окажется достаточным одной такой ошибки, а может быть, пройдет еще некоторое время, пока простой навык изменится в соответствии с новыми обстоятельствами.
Разумеется, для переделки более сложных навыков и привычек, к тому же укоренившихся в силу многолетней тренировки, может потребоваться много времени, но в любом случае большое значение будут иметь индивидуальные особенности высшей нервной деятельности. Выработанные однажды стереотипы не исчезают полностью, они наслаиваются друг на друга, причем более старые системы временных связей затормаживаются новыми и в таком состоянии могут длительно сохраняться. Когда под влиянием каких-либо обстоятельств человек вспоминает давно забытое, это может быть объяснено в некоторых случаях растормаживанием прежде заторможенных временных связей. Последним примером может быть старческая память, при которой в силу возрастных изменений несколько ослабевает тормозной процесс, растормаживаются давно задержанные временные связи и человек начинает отчетливо вспоминать пережитое в детстве, когда-то давно приобретенные сведения, впечатления и т.п.
104
УДК 612(075.8) ББК 28.707.3я73 Ш95
Рецензенты: акад. РАМН, проф. К. В. Судаков (зав. кафедрой нормальной физиологии
ММА им. И. М. Сеченова);
д-р биол. наук, проф. В. В. Раевский (заместитель председателя Ученого совета
Института высшей нервной деятельности и нейрофизиологии РАН);
проф. В. Б. Полянский (зав. лабораторией физиологии сенсорных систем
кафедры высшей нервной деятельности МГУ им. М.В.Ломоносова)
В учебнике приведены классические сведения по проблемам обучения и данные о современном состоянии физиологии головного мозга. Отражены такие вопросы, как роль мотиваций в поведении, типы высшей нервной деятельности, проблема асимметрии полушарий головного мозга человека, участие генома в процессах обучения и памяти и ряд других.
Для студентов биологических специальностей вузов. Может быть полезен студентам психологических факультетов университетов и педагогических вузов.
УДК 612(075.8) ББК 28.707.3я73
© Шульговский В.В., 2003 ISBN 5-7695-0969-4 © Издательский центр «Академия», 2003