

Угасание условного слюноотделительного пищевого рефлекса на шум после предварительной практики короткого (5 с) и длинного (30 с)
отставления (по Д. Е. Соловейчику)
Порядковый |
|
|
|
Слюноотделе- |
|
|
номер приме- |
|
Угасаемый |
Латент- |
ние за 1 30 с, |
<аждые капли |
|
нения |
|
условный |
ный |
(в делениях |
|
|
услов- |
Время |
раздражи-тсль |
период слюн- |
шкалы) |
Примечание |
|
M.U1 U раздра- |
|
(действует |
ной |
|
|
|
жителя |
|
30 с) |
реак- |
|
|
|
без |
|
|
ции |
1 |
2 |
|
подкреп- |
|
|
|
|
|
|
ления |
|
|
|
|
|
|
1 |
9 ч 48 мин |
Шум |
3 |
78 |
58 |
Угасание |
|
|
|
|
|
|
испытывал ось |
2 |
9 ч 51 мин |
То же |
15 |
12 |
3 |
после 31 |
3 |
9 ч 54 мин |
» |
5 |
26 |
14 |
коротко отставленного |
4 |
9 ч 57 мин |
» |
|
0 |
10 |
(5 с) под- |
|
|
|
|
|
|
крепления |
5 |
10 ч 00 мин |
» |
— |
0 |
0 |
|
6 |
10 ч 03 мин |
|
15 |
4 |
34 |
|
7 |
10 ч 06 мин |
» |
— |
0 |
14 |
|
8 |
10 ч 09 мин |
» |
— |
0 |
0 |
|
9 |
10 ч 12 мин |
|
— |
0 |
0 |
|
1 |
9 ч 05 мин |
» |
2 |
80 |
35 |
Угасание |
|
|
|
|
|
|
испытывалось |
2 |
9 ч 08 мин |
» |
25 |
2 |
10 |
после 55 |
|
|
|
|
|
|
длинно |
3 |
9 ч 11 мин |
|
— |
0 |
0 |
отставленных |
4 |
9 ч 14 мин |
» |
— |
0 |
0 |
(30 с) подкреплений |
модействию между собой. Факты суммирования внешнего и внутреннего торможения также, по-видимому, могут указывать, что они имеют одинаковую физиологическую основу.
В чем же заключается физиологический механизм внутреннего вида торможения, вырабатываемого в высших отделах нервной системы?
62
Физиологический механизм условного торможения
Условное торможение является активным процессом — оно вырабатывается. Запаздывательное торможение не является также результатом истощения, так как по мере его выработки величина рефлекса увеличивается.
Физиологи павловской школы интересовались локализацией условного торможения в структурах головного мозга. Хорошо известен факт, что у собаки с удаленной корой больших полушарий угасание ориентировочного рефлекса не происходит. Невозможно у такой собаки получить (выработать) и любой вид внутреннего торможения. Отсюда был сделан вывод, что выработка как условного торможения, так и других временных связей происходит в высших отделах нервной системы, в коре больших полушарий.
В лаборатории И. П. Павлова была предпринята попытка определить, в каком звене условного рефлекса вырабатывается условное торможение. Сложились две точки зрения: согласно первой, условное торможение развивается в корковом конце анализатора условного сигнала, согласно второй — в корковом представительстве безусловного рефлекса. Однако последующие попытки выяснить более точно локализацию условного торможения привели к весьма противоречивым выводам.
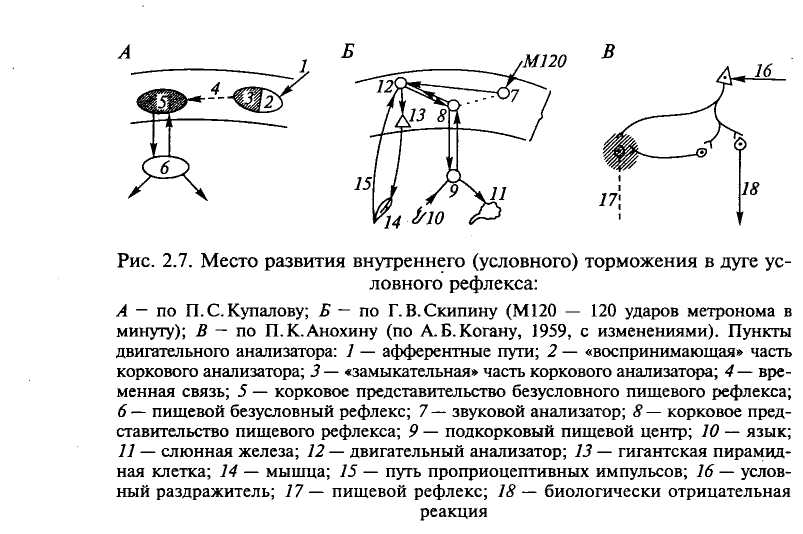
Свойства взаимодействия условных слюнных рефлексов, об разованных путем подкрепления сигнала раздражением выведен ных наружу лоскутов слизистой языка, привели к заключению, что условное торможение развивается как в замыкательной части анализатора сигнального раздражителя, так и в области предста вительства безусловных рефлексов (рис. 2.7, А, В).
Сохранение двигательных компонентов (поднятие лапы) после исчезновения секреторных (слюнных) в процессе угасания услов ного пищевого рефлекса на комплексный раздражитель (метро ном + поднятие лапы) привело к заключению, что условное тор можение возникает не в клетках анализатора сигнального раздра жителя. Наиболее вероятно его первоначальное возникновение где- то на пути к клеткам коркового представительства безусловного пищевого рефлекса, а по мере угасания двигательных компонен тов — в гигантских пирамидных клетках двигательной области (рис. 2.7, Б).
Так как неподкрепление едой вызывает «трудное состояние», имеющее характер целостной отрицательной реакции организма с типичными движениями, вегетативными компонентами (изменение дыхания, сердечной деятельности и др.), было высказано мнение, что условное торможение от неподкрепления разыгрывается в корковом представительстве пищевого рефлекса в результате
63
конкуренции самостоятельного рефлекса «трудного состояния», т.е. по существу торможение, называемое условным, оказывается разновидностью безусловного.
Наконец, так как при наличии нескольких условных рефлексов, выработанных на разные звуковые раздражители при подкреплении разными движениями, угасание одного из них не влияет на подкрепляющее движение, даже если оно сигнализируется совершенно иным звуковым сигналом, было сделано предположение, что условное торможение развивается в ограниченной части двигательного анализатора, связанного с сигнальным раздражителем.
Показатели электрических потенциалов позволяют видеть корреляты1 условного торможения в мозге человека. Если испытуемому дать инструкцию: «При включении лампочки разгибайте кисть», то свет лампочки будет выполнять роль подкрепляющего раздражителя, например, на звуковой сигнал. Теперь звук сам будет вызывать подавление амплитуды биоэлектрических колебаний и увеличение частоты альфа-ритма. Но если перейти к сочетаниям при большом отставлении света от звука, т.е. выработать запаздывание, картина изменяется. Во время своего удлиненного изолиро-
1 Под коррелятами понимают признаки
(например, характер биопотенциалов),
сопровождающие данный физиологический
процесс. По смыслу термин «коррелят»
близок к термину «маркер» в генетике.
Под коррелятами понимают признаки
(например, характер биопотенциалов),
сопровождающие данный физиологический
процесс. По смыслу термин «коррелят»
близок к термину «маркер» в генетике.
64
ванного действия звук начинает усиливать и замедлять колебания альфа-ритма (рис, 2.8). Развивающееся запаздывательное торможение проявило себя перестройкой корковых ритмов: урежением ритмов и устранением быстрых колебаний. Таким образом, наиболее общее изменение в электрических потенциалах мозга при условном торможении — это сдвиг в сторону более медленных ритмов, устранение бета- и замедление альфа-ритма со значительным возрастанием последнего. Такое направление изменений ритмов можно оценить как снижение лабильности — подвижности нервных процессов возбуждения и торможения — нейронов коры.
Термин «лабильность» был введен в начале XX в. в России, в школе Н. Е. Введенского. Лабильность — способность нейронов воспроизводить ритм афферентных возбуждений. При снижении лабильности данной группы нейронов они не способны отвечать на каждый афферентный импульс и впадают в состояние пессиума, что можно интерпретировать как торможение. Состояние нейрона, которое ранее называли пессимальным, хорошо известно и в современной физиологии. Например, по такому механизму тормозятся клетки Пуркинье в коре мозжечка в результате их возбуждения по лиановидным волокнам. Известно, что в ответ на одно возбуждение, приходящее по лиановидному волокну, на мембране клетки Пуркинье развивается высокоамплитудный ВПСП, который приводит к возникновению 2 — 3 потенциалов действия, а затем наступает блокирование натриевой проводимости мембраны (натриевая инактивация), что можно интерпретировать как торможение активности нейрона.
Ввиду отсутствия общепризнанной теории центрального торможения в нервной системе трудно делать какие-либо заключения о природе его частного случая — условного торможения в высших отделах головного мозга.

В соответствии с парабиотеческой гипотезой процесс условного торможения заключается в снижении лабильности нейронов, образующих путь условного рефлекса. Приходящие залпы сигнальных импульсов действуют теперь как пессимальные раздражения и приводят клетки в тормозное состояние. Эта гипотеза основана на гипотезе раздражимости Н. Е. Введенского, в частности на ее положении о переменной лабильности как о факторе перехода клетки от возбудительного к тормозному реагированию.