

4. Пищевые цепи и сети, их состав, формирование и закономерности строения
Функциональными структурными единицами функциональной организации биогеоценоза, как известно, являются пищевые цепи и сети.
Звенья пищевой цепи, составленные сходными по типу питания и образу жизни группами организмов, получили название трофических уровней.
В пищевой цепи обычно выделяются следующие трофические уровни и группы организмов, их составляющие.
I. Зеленые автотрофные растения.
II. Растительноядные животные-фитофаги.
Плотоядные животные-зоофаги (хищники 1-го порядка).
Хищные животные 2-го порядка, обычно более крупные хищники.
Позднее стали выделяться в особую группу (звено) позвоночные и беспозвоночные животные, питающиеся отбросами и трупами животных, — мусорщики.
Начальное звено каждой цепи питания, как видно, представлено популяцией (или популяциями) автотрофного кормового растения, а конечное — наиболее крупным хищником (или хищниками) в данной зоне (белый медведь в Арктике, рысь и бурый медведь в тайге, волк и шакал в аридной полосе, лев, тигр в тропиках и субтропиках). Животные разных трофических уровней так тесно связаны пищевыми отношениями в цепях питания, что стоит произвести какие-либо нарушения в одном звене взаимосвязанных видов живых существ, как произойдет изменение их пищевых взаимоотношений во всем биогеоценозе.
Позднее выяснилось, что некоторые виды животных того или иного трофического уровня одной цепи питания временно или постоянно используют также в качестве источника энергии биомассу живых существ, относящихся к соответствующему трофическому уровню другой цепи питания данного биогеоценоза.
Вследствие такого переплетения пищевыми взаимоотношениями животных разных цепей питания в составе биогеоценоза образуются сложные сочетаний видовых популяций, названные зоологами циклами питания, получившими впоследствии название пищевых сетей.
По мере развития биогеоценотических идей происходило изменение и усложнение представления о пищевых цепях. Цепи питания в трактовке зоологов и зооэкологов представляли собой то, что Дж. Вудвелл (1972) назвал цепью выедания. В такого рода схемах пищевых цепей цикл обмена веществ оставался незавершенным; отсутствовали в схеме и указания на источники материальных и энергетических ресурсов. Поэтому в схеме пищевых цепей Вудвелла параллельно цепи выедания показана цепь разложения, представленная редуцентами, а снизу схемы — источники поступления извне энергии, воды, минеральных элементов и биогенных газов, а пунктирными стрелками — выход отработанной энергии и отходов в сток.
В биологической литературе встречаются следующие определения или краткие фрагментарные характеристики понятия пищевой цепи и сети. По П. Дювиньо и М. Тангу (1968), цепи питания— это «...соподчиненные ряды организмов... Они создают последовательность иерархических уровней созидания и потребления продукции. Переплетаясь, они образуют сложную трофическую (кормовую) сеть».
Ю. Одум (1975) пишет: «Перенос энергии пищи от ее источника — растений — через ряд организмов, происходящий путем поедания одних организмов другими, называется пищевой цепью».
Более развернуто, с учетом более поздних представлений о пищевых цепях, формулирует это понятие Н.С. Абросов с соавторами (1982): «Пути передачи вещества и энергии... через биологические составляющие (экосистему) за счет поглощения внешней, как правило, световой энергии... получили название пищевых цепей; они образуют в природных экосистемах сложную пищевую сеть, обеспечивающую круговорот вещества» (с. 15).
Зарубежными экологами в зависимости от исходного источника энергии различается несколько типов пищевых цепей. Ю. Одум (1968, 1975) выделяет два типа:
I. Пастбищная цепь в составе трофических уровней: 1) продуценты — зеленые растения; 2) первичные консументы — растительноядные животные; 3) вторичные консументы — хищные животные; 4) третичные консументы — вторичные хищники.
II. Детритная цепь: 1) мертвое органическое вещество; 2) микроорганизмы, питающиеся мертвой органикой; 3) детритофаги — потребители детрита; 4) хищники, питающиеся детритофагами.
П. Дювиньо и М. Танг (1968) различают три типа пищевых цепей.
I. Цепи хищников в составе четырех трофических уровней: 1) зеленые кормовые растения; 2) растительноядные животные; 3) мелкие хищники; 4) крупные хищники.
II. Цепи паразитов из двух трофических уровней: 1) живые растения или животные; 2) паразиты на них или в их теле.
III. Цепи сапрофагов также в составе двух трофических уровней: 1) мертвое органическое вещество растительного и животного происхождения; 2) некоторые беспозвоночные животные и микроорганизмы — потребители органики.
Р. Дажо (1975) находит возможным цепь хищников и цепь паразитов слить в одну цепь хищников и паразитов и, в отличие от предыдущих авторов, вводит в ее состав еще один трофический уровень — деструкторы или биоредуценты, состоящий преимущественно из микроорганизмов (бактерий, грибов), разлагающих органические вещества, замыкая тем самым круговорот веществ в пищевой цепи. Выделяемая автором детритная цепь включает два трофических уровня: 1) детрит (мертвое органическое вещество) и 2) консументы — питающиеся детритом организмы (почвенные беспозвоночные животные и микроорганизмы), завершающие разложение органических веществ.
Из отечественных биологов Н.С. Абросов с Б.Г. Ковровым (1977), как и Ю. Одум, различают два типа пищевых цепей:
I. Пастбищная цепь — цепь питания «с передачей энергии от зеленых растений к последующим звеньям цепи путем поедания живых организмов предыдущего звена организмами последующего звена».
II. Детритная цепь — цепь разложения, «которая начинается от мертвого органического вещества, проходит через микроорганизмы, а затем может вновь соединяться с пастбищной цепью».
Анализируя пищевые цепи и сети, зарубежные и отечественные биологи высказывают ряд важных наблюдений и замечаний относительно набора и состава трофических уровней пищевых цепей, функционирования пищевых цепей разного типа, роли и значения разных трофических уровней в функционировании пищевых цепей, сетей и биогеоценозов в целом и пр.
Разнообразие пищевых цепей, по наблюдениям Р. Дажо (1975), связано с разнообразием условий функционирования биокосных систем, в состав которых они входят. Однако чаще, как он полагает, представленные в экосистеме (биогеоценозе) типы пищевых цепей функционируют одновременно, но одна из них при этом доминирует; лишь в особо специфических условиях, например в подземной и абиссальной средах, сохраняется только детритная пищевая цепь.
В природных сообществах к одному трофическому уровню, по словам Ю. Одума (1975), относятся виды организмов, получающие свою пищу от растений «через одинаковое число» звеньев цепи». При этом при каждом переносе энергии с одного трофического уровня пищевой цепи на другой большая часть энергии (до 80-90%) теряется для системы, переходя в тепло. Отсюда «чем короче пищевая цепь (или чем ближе организм к ее началу), — пишет Ю. Одум, — тем больше количество доступной энергии». Вследствие этого ограничивается также и число трофических уровней в пищевой цепи до четырех-пяти. В цепях хищников (пастбищных пищевых цепях) по мере продвижения по цепи животные, по данным Р. Дажо, все более увеличиваются в размерах и уменьшаются численно, а в цепях паразитов, наоборот, животные уменьшаются в размерах, но численность их возрастает.
Из отечественных биологов Н.С. Абросов и Б.Г. Ковров (1977) считают, что каждый трофический уровень пищевой цепи отражает основные черты жизнедеятельности организмов, их место и роль в выполнении определенной функции в переносе веществ и энергии по цепям и в биологическом круговороте веществ всей системы. Энергетическим материалом для функционирования трофические уровней служат биомасса организмов предыдущего трофического уровня или продукты их метаболизма и деструкции отмерших остатков, отбросов и трупов.
Вместе с тем теми же биологами приводятся факты, свидетельствующие о некоторой условности деления пищевых цепей на трофические уровни. Так, Р. Дажо (1975) отмечает, что всеядные животные питаются одновременно растительной и животной пищей, а хищные животные нередко используют в качестве источника энергии значительное количество разнообразных видов жертв. Ю. Одум (1975) утверждает, что популяция одного вида животных может занимать один и более трофических уровней в зависимости от разнообразия пищевого рациона. Н.С. Абросов и Б.Г. Ковров (1977) указывают на принадлежность некоторых видов животных к нескольким трофическим уровням пищевой цепи. Некоторые, в основном растительноядные, виды грызунов, копытных животных могут поедать и животную пищу, а многие растительноядные птицы почти половину своих пищевых потребностей удовлетворяют за счет животной биомассы. Такого рода факты дали повод для критики трофоэнергетической концепции. Тем не менее, «представление о иерархии соподчиненных трофических уровней использования энергии показывает разные стороны энергетической микроструктуры биотической системы и допускает некоторые количественные сопоставления как внутри одного сообщества, так и между разными сообществами».
Известный интерес представляет вопрос о формировании пищевых цепей. Этот процесс весьма рассмотрен П.М. Рафесом (1972), по мнению которого, он протекает как бы поэтапно. Отдельные простые по составу пищевые (трофические) системы, состоящие из двух-трех звеньев, можно наблюдать в природе. Наиболее простую пищевую систему составляет взаимодействие в процессе питания двух особей разных видов. Автором приводятся следующие примеры:
а) листья березы – гусеница березовой пяденицы;
б) гусеница березовой пяденицы – синица.
Такая пищевая система названа биотрофом. При этом в процессе питания, происходит не простая передача вещества и энергии с пищей потребителю, а он (потребитель) перерабатывает ее в собственную биомассу. Из этого следует, что возникшая в процессе такого рода взаимодействия организмов трофическая система является открытой трофоэнергетической системой.
Потребитель того и другого биотрофа представляет собою объект нападения для других видов. В таком случае возникают трофические системы из взаимодействующих особей трех различных видов:
а) листья березы – гусеница березовой пяденицы – синица;
б) гусеница пяденицы – синица – ястреб (хищник). Такого рода трофическая система названа триотрофом.
Рассмотренные триотрофы, как показывает П.М. Рафес, представляют собою отдельные отрезки цепей питания и, следовательно, могут быть объединены в единую полночленную пищевую цепь в следующем составе трофических уровней: листья березы – гусеницы березовой пяденицы – синицы – ястребы.
Однако далеко не всегда процесс формирования пищевых цепей протекает гладко, без каких-либо коллизий. Прослеживая судьбу возникших простых трофических образований — биотрофов и триотрофов, П.М. Рафес констатирует, что организмы взаимодействующих видов находятся в зависимости от наличия корма и от своих естественных врагов. Поэтому судьба потребителя в составе биотрофа или триотрофа может сложиться по-разному. Потребитель (гусеница насекомого или синица) при благополучном стечении обстоятельств может, пройдя соответствующие стадии развития, дать одно или несколько поколений потомства и дожить до естественной смерти, но может и стать объектом паразитизма и испытывать угнетение либо подвергнуться нападению своего врага и быть съеденным, не оставив потомства. Исходя из этого, автор логично приходит к выводу: трофическая связь между двумя индивидуумами (особями) разных видов носит случайный характер, так как успех реализации ее обусловлен множеством разного рода обстоятельств, вследствие чего не всегда обеспечивается при этом существование особи, а следовательно, и продление существования вида. Отсюда и рассмотренные простые трофические системы, основанные на индивидуальных трофических связях, также являются случайными, а потому скоротечными.
Совершенно по-иному складывается судьба трофической системы, как указывает П.М. Рафес, когда в основе ее оказываются не отдельные особи, а видовые популяции, так как при этом не все входящие в их состав, скажем, гусеницы или синицы будут съедены их «врагами»; часть из них достигнет взрослого состояния и доживет до естественной смерти, дав при жизни потомство. Следовательно, трофические системы в таком случае становятся постоянными, долговечными. Этот вывод автора имеет весьма важное, глубоко принципиальное значение, ибо подтверждает, что только видовая популяция как совокупность особей, преодолевая стохастичность случайности, приобретает форму необходимости, вследствие чего становится прочной и долговременной основой состава, строения и функциональной организации биогео-ценотической системы.
При анализе состава и строения цепей питания Ч. Элтоном (1934) выявлен ряд закономерностей, названных им правилами.
1. Правило величины особей, суть которого заключается в том, что цепи питания, как правило, идут от более мелких форм к более крупным. Однако, поскольку размеры животных не могут увеличиваться беспредельно, число звеньев цепи не может превышать пяти-шести. Правда, из этого правила имеются исключения, приведенные Р. Дажо (1975): например, нападение волков на более крупных животных, чем сами, благодаря охоте сообща или умерщвление змеями крупных животных посредством выделяемого яда и пр.
2. Правило пирамиды чисел, сформулированное так: число особей в каждом предыдущем звене бывает значительно больше, чем в последующем. Только при этом условии смертность видов-жертв от истребления хищниками будет компенсироваться и численность их будет поддерживаться на определенном уровне. Пирамида чисел, как отмечает Р. Дажо, тем выше, чем большее число уровней входит в состав данной пищевой цепи. Он приводит и некоторые исключения, например пищевые цепи паразитов, а также сапрофагов.
3. Правило биомассы, вытекающее из предыдущего правила, заключается в том, что биомасса с каждым последующим звеном цепи уменьшается в среднем в отношении 10:1, так как далеко не вся масса какого-либо звена превращается в массу последующего звена, часть ее расходуется на процессы жизнедеятельности организмов, на их воспроизводство и т. п. Исключение из этого правила относится к водным биокосным системам, в которых, например, биомасса фитопланктона меньше биомассы питающегося ею зоопланктона; правда, она значительно быстрее возобновляется.
Р. Дажо (1975) приводит еще одно правило:
4) правило пирамиды энергии, где пирамида имеет вид треугольника с вершиной, обращенной вверх, в связи с потерей энергии при переходе от одного трофического уровня к другому. Это правило наилучшим образом выражает трофоэнергетическую суть биокосной системы, но для этого в настоящее время часто не хватает необходимых данных.
Итак, что же представляют собою пищевые цепи и трофические уровни, их составляющие? Ю. Одум (1975) подчеркивает, что «трофическая классификация делит на группы не сами виды, а их типы жизнедеятельности» (с. 85), а Н.С. Абросов и Б.Г. Ковров (1977) утверждают, что «трофические уровни пищевых цепей — это не что иное, как живые компоненты биогеоценоза» (с. 10).
Из всего изложенного следует, что взаимодействующими «субъектами», образующими трофические уровни в пищевых цепях, несомненно, являются видовые популяции (ценопопуляции) живых существ, вступающие приовместном обитании в составе ценоза в сложные трофические (пищевые) и другие взаимоотношения между собой.
Трофические уровни (звенья) в составе пастбищной цепи (цепи выедания, по Вудвеллу) формируются из ценопопуляции автотрофного ценокомплекса (I уровень) и из ценопопуляции биотрофного ценокомплекса (II уровень фитофагов, III и IV — уровни зоофагов 1-го и 2-го порядка); источником энергии тех и других служат воплощенная в последовательно трансформируемой биомассе световая энергия солнечной радиации и материальные пищевые и другие ресурсы местообитания. Трофический уровень детритной цепи (цепи разложения, по Вудвеллу) состоит из ценопопуляции сапротрофного ценокомплекса, обитающих как в наземной (воздушной), так и в почвенной среде биотопа, и в качестве источника энергии используют отмершие остатки, отбросы и мертвые органические вещества растительного и животного происхождения, осуществляя одновременно и синтез гумусовых и других веществ.
Следовательно, живые компоненты биокосных систем, представленные в природе ценокомплексами автотрофных ценопопуляций, биотрофных ценопопуляций (фитофаги и зоофаги) и ценопопуляций сапротрофов, нельзя отождествлять с трофическими уровнями пищевых цепей, как нельзя и обезличивать сами трофические уровни. Трофические уровни — это формы, в которые облекаются в ходе функционирования в составе пищевой цепи видовые ценопопуляции соответствующих ценокомплексов биогеоценотических систем. При этом не следует забывать, что в состав биогеоценоза входит не одна, а целый ряд пищевых цепей, базирующихся на ценопопуляциях автотрофных кормовых ценозообразователей. Пищевые же цепи (и сети) того или иного типа, следовательно, должны рассматриваться как важнейшие функциональные структурные единицы функциональной организации биоценоза и биогеоценоза в целом, весьма сложные по популяционно-видовому составу живых существ и по выполняемой в биогеоценозе функции.
Хищные
птицы (лунь
и др.)
Наряду с пищевыми цепями и сетями, биологами выявлена еще одна функциональная структурная единица функциональной организации биогеоценоза — это так называемая консорция. Проблема консорции — многоплановая проблема, свидетельством чему служит тот факт, что это понятие и термин почти одновременно независимо друг от друга были предложены зоологом В.Н. Беклемишевым (1951) и ботаником Л.Г. Раменским (1952). Л.Г. Раменский дал такое, несколько вольное, определение этого понятия: консорции — это «сочетания разнородных организмов, тесно связанных друг с другом в их жизнедеятельности известной общностью их судьбы» (с. 186). Он приводит примеры, иллюстрирующие это понятие. Одна из консорций елового сообщества, представленная популяцией ели и целым рядом популяций других видов, тесно связанных с нею в своей жизнедеятельности: а) микоризные грибы — на ее корнях; б) кустистый лишайник — на ветвях; в) корковые лишайники — на коре стволов; г) жуки-короеды и усачи — в стволах; д) еловая огневка— в кроне деревьев; е) бурундуки и белки, питающиеся семенами ели и пр. (рис. 18). Консорция дубового сообщества, состоящая из популяции дуба и связанных с ним растительных и животных паразитов, эпифитов, симбионтов, вредителей из насекомых и пр.
Таким образом, центром каждой консорции служит популяция какого-либо высшего автотрофного растения, с которым тесно связаны трофическими, симбиотическими, паразитическими и другими отношениями разнообразные виды растений, преимущественно низших, позвоночных и беспозвоночных животных и микроорганизмов. Эти биологические виды-партнеры названы консортами, а связи и отношения консортов с популяцией автотрофного кормового растения — консортивными связями.
В отличие от Л.Г. Раменского, В.Н. Беклемишев предложенный им термин употребил в мужском роде — «консорций». По его мнению, организмы входят в состав биогеоценоза не сами по себе, а в составе какого-либо консорция (или иначе — консорции). Консорция состоит из особи-эдификатора и целого ряда поселяющихся на теле или в теле эдификатора особей других видов. Автором приводятся следующие примеры консорций: а) сосна с микоризными грибами, мхами, насекомыми и пр.; б) полевка с эктопаразитами, гельминтами, простейшими, бактериями и т. п.
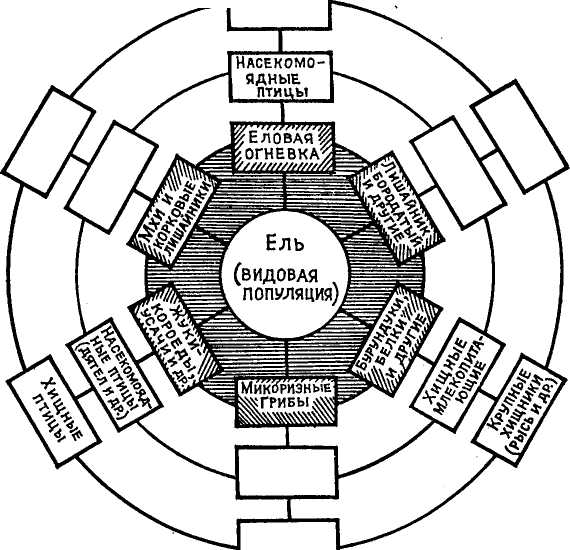
Схема консорции популяции ели (по данным Л. Г. Раменского, 1952, и В. В. Мазинга, 1966)
Следовательно, центром консорции, по Беклемишеву, является не популяция вида как поселение особей, а одна особь какого-либо доминирующего вида, причем это может быть как растение, так и животное (полевка в данном случае).
В дальнейшем понятие консорции значительно расширилось. В.В. Мазинг (1966) предложил в состав консорции включать как виды организмов, непосредственно связанные с популяцией автотрофного кормового растения, образующие I круг (концентр) вокруг нее, так и виды, связанные с нею опосредованно через организмы I концентра, составляющие II, III и IV концентры. В результате консорция выглядит весьма сложной как по составу, так и по своему строению единицей функциональной организации биоценоза и биогеоценоза в целом (см. рис. 18).
Любая консорция, как видно на рис. 18, состоит из центрального организма или видовой популяции, называемых то ядром, то эдификатором или детерминантом консорции, и группы организмов-консортов, связанных в своей жизнедеятельности с центральным видом. Прежде всего, что считать центром консорции? Е.М. Лавренко (1959), вслед за Л.Г. Раменским, предлагает центром консорции в сухопутных системах считать популяцию высшего автотрофного зеленого растения, являющегося как бы «энергетической установкой биоценоза». По Т.А. Работнову (1969), центральным видом, детерминантом консорции могут быть не только высшие, но всякие автотрофные растения, на суше, в частности, мхи, водоросли и др. А. А. Корчагин (1977), в согласии с В.Н. Беклемишевым, находит, что на положении центра, ядра консорции может оказаться не только автотрофное растение, но и гетеротрофное животное. И.А. Селиванов (1974) и П.М. Рафес (1970) идут еще дальше, считая, что в качестве центра консорции следует признать не только живые авто- и гетеротрофные существа, но и мертвые тела (пни, валежник, трупы животных). Точка зрения последних авторов разделяется и Дылисом (1978), но консорции, развивающиеся на мертвых (отмерших) телах, по его мнению, должны быть выделены в особую группу. Но Б.А. Быков (1970) высказывается против включения в состав консорции организмов, использующих в качестве источника энергии разлагающиеся ткани мертвых растений и животных.
Еще более глубокие расхождения во взглядах биологов обнаружились по вопросу о составе консортов и в связи с этим о структуре и объеме самих консорций. К группе консортов принято относить: а) потребителей живой биомассы — растительной, животной — это так называемые биотрофы (консументы) и б) потребителей отмерших остатков и мертвого органического вещества растительного и животного происхождения — это сапротрофы (биоредуценты). Т.А. Работновым сюда же относятся: в) эккрисотрофы, использующие в качестве источника энергии прижизненные выделения организмов, и г) эпифиты, для которых живые существа, образующие ядро консорции, служат лишь субстратом для прикрепления. Иными словами, в состав консортов входят организмы или видовые популяции, связанные с детерминантом консорции трофически или топически.
В состав I концентра консорции усложненной структуры входят консорты, непосредственно связанные с центральным видом консорции. Одни из них связаны с ним трофически, получая от него вещества и энергию: а) животные-фитофаги, большей частью беспозвоночные — симбионты, паразиты, вредители, в том числе насекомые (жуки, чешуекрылые, перепончатокрылые); б) фитопаразиты (бактерии, грибы, актиномицеты, вирусы и др.); в) растения-полупаразиты, получающие от центрального вида консорции только вещества; г) азотфиксирующие растения-симбионты, получающие от центрального вида энергию и вещества (за исключением азота). Другие консорты связаны с центральным видом трофически и топически: а) растения-паразиты; б) животные, поселяющиеся внутри его особей или вступающие с ними в симбиотические отношения. У третьих консортов связь с детерминантом консорции только топическая: а) растения-эпифиты (мхи, лишайники, некоторые высшие растения), использующие его в качестве субстрата для прикрепления; б) животные, которые находят здесь убежище для себя и для своего потомства.
II концентр консорции составляют видовые популяции, организмы которых в качестве источника энергии используют биомассу организмов I концентра как в живом, так и в отмершем состоянии, а также их экскременты. Сюда относятся: а) животные-зоофаги (хищники 1-го порядка) — млекопитающие, птицы, некоторые насекомые (жужелицы, божьи коровки, пауки и пр.); б) зоопаразиты животных-фитофагов (личинки насекомых, кровососущие насекомые); в) беспозвоночные животные-сапрофаги, некрофаги, копрофаги, живущие за счет мертвого органического вещества и экскрементов животных.
В состав III и последующих концентров консорции входят видовые популяции живых существ, использующих в процессах своей жизнедеятельности энергию, заключенную в биомассе организмов предшествующих концентров: а) животные-зоофаги (хищники 2-го и 3-го порядка); б) зоопаразиты плотоядных животных предшествующих концентров, в) животные-сапрофаги и микроорганизмы, живущие за счет использования энергии мертвого органического вещества (насекомые, бактерии и пр.).
При определении места, позиции, занимаемой той или иной ценопопуляцией в составе консорции, выявился ряд трудностей. Так, мхи и лишайники обычно относятся к консортам, но на моховых болотах и, очевидно, в некоторых биогеоценозах тундры они становятся основными продуцентами, а потому, как считает А. А. Корчагин (1977), должны рассматриваться в таком случае как центральные виды консорций. Автотрофные полупаразиты, которые обычно относятся к консортам-консументам, по мнению Т.А. Работнова, следует считать консортами-эпифитами, поскольку они получают от детерминанта лишь воду и элементы минерального питания; последний же выступает в таком случае как субстрат для их прикрепления. Насекомоядные хлорофиллоносные растения являются продуцентами и одновременно зоофагами-консументами, получающими от своих жертв только азот и фосфор. Поэтому Т.А. Работнов склонен относить их к числу центральных растений соответствующих консорций. Лианы, особенно деревянистые, нередко настолько угнетают растение-опору, что становятся основными продуцентами консорции. Следовательно, вопрос о позиции лианы в составе консорции, по мнению А.А. Корчагина, должен решаться по-разному, в зависимости от степени угнетения опоры-хозяина и от доли участия каждого из них в продуцировании органики.
Значение и доля участия консортов в потреблении и трансформации органических веществ в составе консорции неодинаковы. Из растительных консортов наибольшее значение в этом, по словам А. А. Корчагина (1977), имеют почвенные бактерии и грибы; из наземных растений — мхи, лишайники и водоросли, а также паразиты и полупаразиты — из высших растений. Из животных видов к числу наиболее важных консортов относятся беспозвоночные, особенно насекомые, среди них фитофаги и фитопаразиты.
Для характеристики консорции в целом наибольшее значение, как утверждает В.В. Мазинг (1966), имеет I концентр консорции, состоящей из видов, непосредственно связанных с автотрофным детерминантом. «Чем дальше он от центра, тем меньше специфичных для консорции и облигатных связей. Виды высших звеньев пищевых цепей («хищники хищников») требуют большего количества пищи, поступающего не только из других консорций, но и из различных биоценозов, часто пространственно разобщенных».
Обращает на себя внимание исключительное видовое богатство консортов большинства автотрофных ценозообразователей, в особенности древесных и кустарниковых пород. Так, по данным В. В. Мазинга (1966), численный состав консортов только I концентра болотной формы сосны на верховых болотах Эстонии составляет не менее 100 видов, в том числе: грибов (аскомицетов, базиомицетов и несовершенных)—30 видов, эпифитных мхов и лишайников — 32 вида, насекомых-фитофагов (жуков, чешуекрылых, перепончатокрылых и др.) —38 видов. Общее число консортов I концентра пушистой и бородавчатой березы, по данным того же автора (1976), достигает почти 1000 видов, в частности: грибов (паразитов и симбионтов) — 147 видов, эпифитных мхов и лишайников— 69, высших цветковых растений-паразитов— 1, животных-консументов (паукообразных, насекомых и других беспозвоночных) — 636, птиц (куриных, воробьиных, дятловых) — 8, млекопитающих (грызунов, парнокопытных) — 12; всего 974 вида, не считая водорослей и других низших организмов! Среди консортов беспозвоночных животных особенно много листогрызущих форм, а также стволовых и подкоровых обитателей. Менее богаты видами-консортами, по сравнению с деревьями и кустарниками, травянистые цветковые детерминанты. К числу бедных по видовому составу относятся консорции плаунов, хвощей и папоротникообразных ценозообразователей. Видовое богатство консорций, по словам В.В. Мазинга (1976), определяется: а) филогенетическим возрастом детерминанта и динамикой его ареала; б) разнообразием поставляемого детерминантом корма для консортов и экологических ниш для их обитания.
В связи с изменившимся представлением о консорции по сравнению с первоначальным появился целый ряд новых определений консорции отечественными биологами: Е.М. Лавренко (1959), В.В. Мазинга (1966), Т.А. Работнова (1969), Б.А. Быкова (1970), А.Г. Воронова (1976), Н.В. Дылиса (1974 и 1978) и др. В этих определениях консорции подчеркиваются следующие присущие ей черты: а) центром, «ядром» консорции большинством биологов признается видовая популяция, а не отдельно взятый организм, причем популяция автотрофного вида (или, по Мазингу, также группы экологически близких видов) растений, а из автотрофов — популяция самостоятельно существующего вида (т.е. не эпифита и не полупаразита); б) объединение или сочетание центральной видовой популяции происходит с организмами всех входящих в состав консорции видов, как растительных, так и животных, как высших, так и низших; в) в основе такого рода сочетания видовых популяций консорции лежат трофические или топические связи и отношения или те и другие одновременно; г) сформировавшаяся таким образом консорция как совокупность разнообразных видовых популяций большинством биологов рассматривается как единица функциональной организации биогеоценоза, а не фитоценоза, как считалось ранее.
Из всех современных определений консорции названных авторов наиболее полным является определение Т.А. Работнова (1969). «...Под консорцией следует понимать сочетание популяции любого самостоятельно существующего (т.е. не эпифита) в пределах определенного фитоценоза автотрофного растения и совокупности организмов, связанных с ними трофически (консументов, редуцентов), или использующих как субстрат, для прикрепления (эпифиты), или как источник воды и элементов минерального питания (автотрофные полупаразиты)».
Однако наиболее точно раскрывающим суть консорций как функциональных структурных единиц биогеоценоза следует признать определение их Н.В. Дылисом (1978): «С биогеоценологической точки зрения, консорции есть функциональные структуры биоты, отражающие в своей совокупности разнообразие путей перемещения и трансформации веществ и энергии от первичных продуцентов биогеоценоза ко всем в нем участвующим консументам и деструкторам».