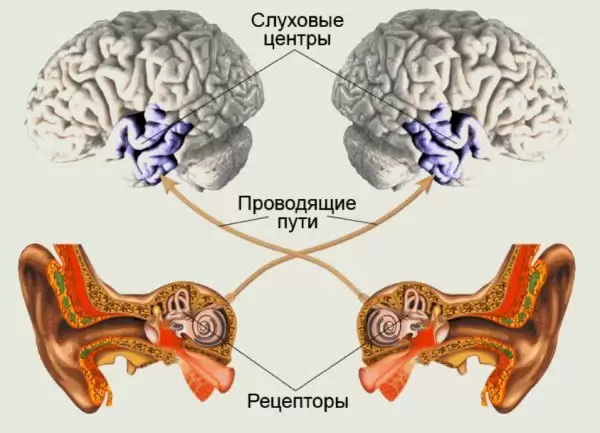
Слуховой анализатор. Механизм восприятия звуков разной частоты.
Слуховой анализатор воспринимает колебания воздуха и трансформирует механическую энергию этих колебаний в импульсы, которые в коре головного мозга воспринимаются как звуковые ощущения.
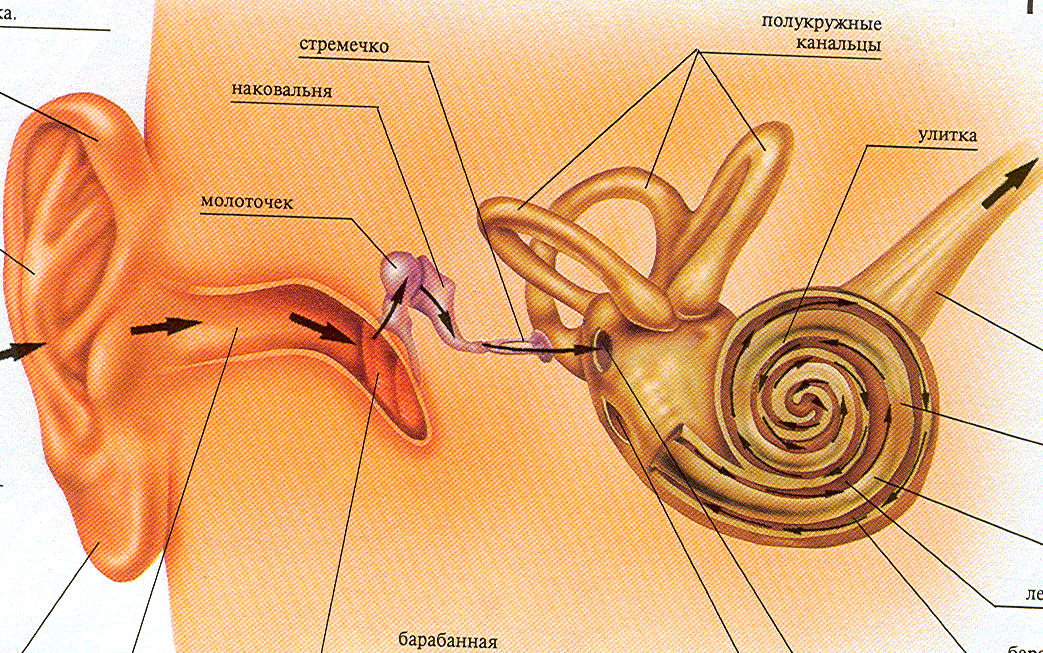
Воспринимающая часть слухового анализатора включает - наружное, среднее и внутреннее ухо (рис. 11.8.). Наружное ухо представлена ушной раковиной (звукоуловитель) и наружным слуховым проходом, длина которого составляет 21-27 мм, а диаметр 6-8 мм. Наружное и среднее ухо разделяет барабанная перепонка - мало податливая и слабо растягивающаяся мембрана.
Среднее ухо состоит из цепи соединенных между собой косточек: молоточек, наковальня и стремечко. Рукоятка молоточка прикрепляется к барабанной перепонке, основание стремечка - к овальному окну. Это своеобразный усилитель который в 20 раз усиливает колебания. В среднем ухе, кроме того, имеется две маленькие мышцы, прикрепляющиеся к косточкам. Сокращение этих мышц приводит к уменьшению колебаний. Давление в среднем ухе выравнивается за счет евстахиевой трубы, которая открывается в ротовую полость.
Внутреннее ухо соединено со средним при помощи овального окна, к которому прикрепляется стремечко. Во внутреннем ухе находится рецепторный аппарат двух анализаторов - воспринимающего и слухового (рис. 11.9.). Рецепторный аппарат слуха представлен улиткой. Улитка, длиной 35 мм и имеющая 2,5 завитка, состоит из костной и перепончатой части. Костная часть разделена двумя мембранами: основной и вестибулярной (рейснеровой) на три канала (верхний - вестибулярный, нижний - тимпанический, средний - барабанный). Средняя часть, называется улиточный ход (перепончатый). У верхушки - верхние и нижние каналы связаны геликотремой. Верхние и нижние каналы улитки заполнены перилимфой, средние - эндолимфой. Перилимфа по ионному составу напоминает плазму, эндолимфа - внутриклеточную жидкость (в 100 раз больше ионов К и в 10 раз ионов Nа).
Основная мембрана состоит из слабо натянутых эластических волокон, поэтому может колебаться. На основной мембране - в среднем канале расположены звуковоспринимающие рецепторы - кортиев орган (4 ряда волосковых клеток - 1 внутренний (3,5 тыс. клеток) и 3 наружных - 25-30 тыс. клеток). Сверху - тектореальная мембрана.
Механизмы проведения звуковых колебаний. Звуковые волны пройдя через наружный слуховой проход колеблют барабанную перепонку, последняя приводит в движение косточки и мембрану овального окна. Колеблется перилимфа и к вершине колебания затухают. Колебания перилимфы передаются на вестибулярную мембрану, а последняя начинает колебать эндолимфу и основную мембрану.
В улитке регистрируется: 1) Суммарный потенциал (между кортиевым органом и средним каналом - 150 мВ). Он не связан с проведением звуковых колебаний. Он обусловлен уравнем окислительно-восстановительных процессов. 2) Потенциал действия слухового нерва. В физиологии также известен и третий - микрофонный - эффект заключающий в следующем: если в улитку ввести электроды и соединить с микрофоном, предварительно усилив его, и произносить в ухо кошке различные слова, то микрофон воспроизводит эти же слова. Микрофонный эффект генерируется поверхностью волосковых клеток, т. к. деформация волосков приводит к появлению разности потенциалов. Однако, этот эффект превосходит энергию вызвавших его звуковых колебаний. Отсюда микрофонный потенциал - непростое преобразование механической энергии в электрическую, а связан с обменными процессами в волосковых клетках. Местом возникновения микрофонного потенциала является область корешков волосков волосковых клеток. Звуковые колебания, действующие на внутреннее ухо, накладывают возникающий микрофонный эффект на эндокохлеарный потенциал.
Суммарный потенциал отличается от микрофонного тем, что отражает не форму звуковой волны, а ее огибающую и возникает при действии на ухо высокочастотных звуков (рис. 11.10.).
Потенциал действия слухового нерва генерируется в результате электрического возбуждения, возникающего в волосковых клетках в виде микрофонного эффекта и суммарного потенциала.
Между волосковыми клетками и нервными окончаниями имеются синапсы, при этом имеет место и химический и электрический механизмы передачи.
Механизм передачи звука различной частоты. В течение длительного времени в физиологии господствовала резонаторная теория Гельмгольца: на основной мембране натянуты струны различной длины, подобно арфе они имеют разную частоту колебаний. При действии звука начинает колебаться та часть мембраны, которая настроена в резонанс данной частоте. Колебания натянутых нитей раздражают соответствующие рецепторы. Однако, эта теория критикуется, т. к. струны не натянуты и их колебания в каждый данный момент включают слишком много волокон мембраны.
Заслуживает внимания теория Бекеше. В улитке имеется явление резонанса, однако, резонирующим субстратом являются не волокна основной мембраны, а столб жидкости определенной длины. По данным Бекеше, чем больше частота звука, тем меньше длина колеблющегося столба жидкости. При действии звуков низкой частоты длина колеблющегося столба жидкости увеличивается, захватывая большую часть основной мембраны, причем колеблются не отдельные волокна, а значительная их часть. Каждой высоте тона соответствует определенное количество рецепторов.
В настоящее время наиболее распространенной теорией восприятия звука разной частоты является “теория места”, согласно которой не исключается участие воспринимающих клеток в анализе слуховых сигналов. Предполагается что волосковые клетки, расположенные на различных участках основной мембраны обладают различной лабильностью, что оказывает влияние на звуковые восприятия, т. е. речь идет о настройке волосковых клеток на звуки разной частоты.
Повреждения в различных участках основной мембраны приводит к ослаблению электрических явлений, возникающих при раздражении звуков разной частоты.
Согласно резонансной теории, различные участки основной пластинки реагируют колебанием своих волокон на звуки разной высоты. Сила звука зависит от величины колебаний звуковых волн, которые воспринимаются барабанной перепонкой . Звук будет тем сильнее , чем больше величина колебаний звуковых волн и соответственно барабанной перепонки , Высота звука зависит от частоты колебаний звуковых волн, Большая частота колебаний в единицу времени будет. восприниматься органом слуха в виде более высоких тонов ( тонкие, высокие звуки голоса ) Меньшая частота колебаний звуковых волн воспринимается органом слуха в виде низких тонов ( басистые, грубые звуки и голоса ) .
Восприятие высоты, силы звука и локализации источника звука начинается с попадания звуковых волн в наружное ухо, где они приводят в движение барабанную перепонку. Колебания барабанной перепонки через систему слуховых косточек среднего уха передаются на мембрану овального окна, что вызывает колебание перилимфы вестибулярной (верхней) лестницы. Эти колебания через геликотрему передаются перилимфе барабанной (нижней) лестницы и доходят до круглого окна, смещая его мембрану по направлению к полости среднего уха. Колебания перилимфы передаются также на эндолимфу перепончатого (среднего) канала, что приводит в колебательные движения основную мембрану, состоящую из отдельных волокон, натянутых, как струны рояля. При действии звука волокна мембраны приходят в колебательные движения вместе с рецепторны-ми клетками кортиева органа, расположенными на них. При этом волоски рецепторных клеток контактируют с текториальной мембраной, реснички волосковых клеток деформируются. Возникает вначале рецепторный потенциал, а затем потенциал действия (нервный импульс), который далее проводится по слуховому нерву и передается в другие отделы слухового анализатора.
Электрические явления в улитке.
При отведении электрических потенциалов от разных частей улитки обнаружено пять различных феноменов: два из них — мембранный потенциал слуховой рецепторной клетки и потенциал эндолимфы — не обусловлены действием звука; три электрических явления — микрофонный потенциал улитки, суммационный потенциал и потенциалы слухового нерва— возникают под влиянием звуковых раздражений.
1. Мембранный потенциал слуховой рецепторной клетки характеризует состояние покоя.
2.Потенциал эндолимфы, или эндокохлеарный потенциал, обусловлен различным уровнем окислительно-восстановительных процессов в каналах улитки, в результате чего возникает разность потенциалов (80 мВ) между перилимфой среднего канала улитки (потенциал которой имеет положительный заряд) и содержимым верхнего и нижнего каналов. Этот эндокохлеарный потенциал оказывает влияние на мембранный потенциал слуховых рецепторных клеток, создавая у них критический уровень поляризации, при котором незначительное механическое воздействие во время контакта волосковых рецепторных клеток с текториальной мембраной приводит к возникновению в них возбуждения.
3. Микрофонный эффект: Если ввести в улитку электроды, соединить их с динамиком через усилитель и подействовать на ухо звуком, то динамик точно воспроизведет этот звук. Описываемое явление называют микрофонным эффектом улитки, а регистрируемый электрический потенциал назван кохлеарным микрофонным потенциалом. Доказано, что он генерируется на мембране волосковой клетки в результате деформации волосков. Частота микрофонных потенциалов соответствует частоте звуковых колебаний, а амплитуда потенциалов в определенных границах пропорциональна интенсивности звука. Потенциал генерируется на мембране волосковой клетки в результате деформации волосков при соприкосновении с текториальной мембраной. Частота микрофонных потенциалов соответствует частоте звуковых колебаний, а амплитуда потенциалов в определенных границах пропорциональна интенсивности звуков речи. Звуковые колебания, действующие на внутреннее ухо, приводят к тому, что возникаюший микрофонный эффект накладывается на эндокохлеарный потенциал и вызывает его модуляцию.
4. В ответ на сильные звуки большой частоты (высокие тона) отмечают стойкий сдвиг исходной разности потенциалов. Это явление получило название суммационного потенциала. Различают положительный и отрицательный суммационные потенциалы. Их величины пропорциональны интенсивности звукового давления и силе прижатия волосков рецепторных клеток к покровной мембране. Суммационный потенциал отличается от микрофонного потенциала тем, что отражает не форму звуковой волны, а ее огибающую. Он представляет собой совокупность микрофонных потенциалов, возникающих при действии сильных звуков с частотой ныше 4000 — 5000 Гц.
Микрофонный и суммационный потенциалы рассматривают как суммарные рецепторные потенциалы волосковых клеток. Имеются указания, что отрицательный суммационный потенциал генерируется внутренними, а микрофонный и положительный суммационные потенциалы — наружными волосковыми клетками.
И наконец, в результате возбуждения рецепторов происходит генерация импульсного сигнала в волокнах слухового нерва.
5. Электрическая активность путей и центров слуховой системы. Даже в тишине по волокнам слухового нерва следуют спонтанные импульсы со сравнительно высокой частотой (до 100 в секунду). При звуковом раздражении частота импульсации в волокнах нарастает и остается повышенной в течение всего времени, пока действует звук. Степень учащения разрядов различна у разных волокон и обусловлена интенсивностью и частотой звукового воздействия (см. рис. 14.14). В центральных отделах слуховой системы много нейронов, возбуждение которых длится в течение всего времени действия звука. На низких уровнях слуховой системы сравнительно немного нейронов, отвечающих лишь на включение и выключение звука (нейроны on-, off- и on-off- типа). На высоких уровнях системы процент таких нейронов возрастает. В слуховой зоне коры большого мозга много нейронов, вызванные разряды которых длятся десятки секунд после прекращения звука.
Потенциал действия слухового нерва регистрируется в его волокнах, частота импульсов соответствует частоте звуковых волн, если она не превышает 1000 Гц. При действии более высоких тонов частота импульсов в нервных волокнах не возрастает, так как 1000 имп/с — это почти максимально возможная частота генерации импульсов в волокнах слухового нерва. Потенциал действия в нервных окончаниях регистрируется через 0,5 — 1,0 мс после возникновения микрофонного эффекта, что свидетельствует о си-наптической передаче возбуждения с волосковой клетки на волокно слухового нерва.
Резонансная теория Гельмгольца основывалась на предположении, что отдельные волокна основной мембраны настроены, как струны, на различные звуковые частоты.
Восприятие звуков различной высоты, обусловлено тем, что каждое волокно основной мембраны настроено на звук определенной частоты.
Как известно, концевым аппаратом слухового нерва является орган Корти, покоящийся на основной перепонке, идущей вдоль всего спирального костного канала, называемого улиткой. Основная перепонка состоит из большого количества (около 24 000) поперечных волокон, длина которых постепенно уменьшается от вершины улитки к ее основанию. По резонансной теории Гельмгольца, каждое такое волокно настроено, подобно струне, на определенную частоту колебаний. Когда до улитки доходят звуковые колебания определенной частоты, то резонирует определенная группа волокон основной перепонки и возбуждаются только те клетки органа Корти, которые покоятся па этих волокнах. Звуки низкой частоты воспринимаются длинными волнами основной мембраны, расположенными ближе к верхушке улитки, звуки высокой частоты воспринимаются короткими волокнами основной мембраны, расположенными ближе к основанию улитки. При действии сложного звука возникают колебания различных волокон мембраны.
В современной интерпретации резонансный механизм лежит в основе теории места, в соответствии с которой в состояние колебания вступает вся мембрана. Однако максимальное отклонение основной мембраны улитки происходит только в определенном месте. При увеличении частоты звуковых колебаний максимальное отклонение основной мембраны смещается к основанию улитки, где располагаются более короткие волокна основной мембраны, — у коротких волокон возможна более высокая частота колебаний. Возбуждение волосковых клеток именно этого участка мембраны при посредстве медиатора передается на волокна слухового нерва в виде определенного числа импульсов, частота следования которых ниже частоты звуковых волн (лабильность нервных волокон не превышает 800 — 1000 Гц). Частота воспринимаемых звуковых волн достигает 20 000 Гц. Таким способом осуществляется пространственный тип кодирования высоты и частоты звуковых сигналов.
Отдельные нейроны на разных уровнях слуховой сенсорной системы настроены на определенную частоту звука, т.е. каждый нейрон имеет свой специфический частотный порог, свою определенную частоту звука, на которую реакция нейрона максимальна. Таким образом, каждый нейрон из всей совокупности звуков воспринимает лишь определенные достаточно узкие участки частотного диапазона, не совпадающие между собой, а совокупности нейронов воспринимают весь частотный диапазон слышимых звуков, что и обеспечивает полноценное слуховое восприятие