

Контрольные вопросы
Из каких частей состоит нервно-мышечный синапс?
Какой медиатор используется для передачи ПД в мионевральном синапсе?
Какие рецепторы находятся на постсинаптической мембране нервно-мышечного синапса?
Какую функцию выполняет фермент ацетилхолинэстераза в синапсе?
Из каких этапов состоит нервно-мышечная передача возбуждения?
Каким образом можно воздействовать на синапс фармакологическими препаратами?
1.5. Механизм мышечного сокращения
В процессе сокращения мышечного волокна в нем происходят следующие преобразования:
А. Электрохимическое преобразование (рис. 9) - это последовательность процессов, в результате которых потенциал действия плазматической мембраны мышечного волокна приводит к запуску цикла поперечных мостиков.

Рис. 9. Распространение потенциала действия по сарколемме мышечного волокна и выброс ионов кальция из цистерн саркоплазматического ретикулума
Выделяют следующие этапы:
1. Генерация ПД.
2. Распространение ПД по Т-системе.
3. Электрическая стимуляция зоны контакта Т-системы и саркоплазматического ретикулума, активация ферментов, образование инозитолтрифосфата, повышение внутриклеточной концентрации ионов Са2+.
Б. Хемомеханическое преобразование:
В 1954 г. H.T. Huxley и A.F. Huxley независимо друг от друга для объяснения мышечного сокращения сформулировали теорию скольжения нитей. Авторы данной теории утверждают, что укорочение саркомера, а следовательно, и мышцы в момент сокращения происходит в результате активного скольжения актиновых нитей относительно миозиновых нитей. При этом саркомер способен укоротиться на 20-30% своей длины. Основные положения этой теории: а) ионы Ca2+ запускают механизм мышечного сокращения; б) за счет ионов Ca2+ происходит скольжение тонких актиновых нитей по отношению к миозиновым.
4. Взаимодействие ионов Са2+ с тропонином, освобождение активных центров на актиновых филаментах.
5. Взаимодействие миозиновой головки с актином, вращение головки и развитие эластической тяги.
6. Скольжение нитей актина и миозина относительно друг друга, уменьшение размера саркомера, развитие напряжения или укорочение мышечного волокна.
В покое взаимодействие тонких и толстых нитей невозможно, т.к. миозин-связывающие участки актина заблокированы тропомиозином (рис. 10).
При высокой концентрации Ca2+ эти ионы связываются с тропонином C и вызывают конформационные изменения тропомиозина, приводящие к разблокированию миозин-связывающих участков. От концентрации Ca2+ в цитоплазме зависит количество занятых Ca2+-связывающих участков тропонина, что в свою очередь, определяет, какое количество участков актина доступно для взаимодействия с поперечными мостиками.
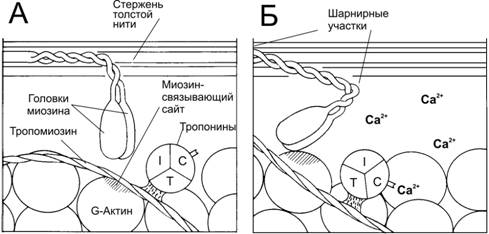
Рис. 10. Ca2+–зависимый механизм регуляции взаимодействия актина с миозином
Головки миозина, несущие продукты гидролиза АТФ (АДФ + Фн), присоединяются к тонкой нити и изменяют свою конформацию, создавая тянущее усилие: — тонкие нити начинают скользить между толстыми. За счёт шарнирного участка в области шейки миозина происходит гребковое движение, продвигающее тонкую нить к центру сaркомера. В результате происходит скольжение тонких нитей относительно толстых. Затем головка миозина связывается с молекулой АТФ, что приводит к отделению миозина от актина. Последующий гидролиз АТФ восстанавливает конформированную молекулу миозина, готовую вступить в новый цикл.
Каждый цикл взаимодействия тонких и толстых нитей имеет несколько стадий (рис. 11). Последовательность событий, начиная от связывания поперечного мостика с тонким филаментом и до момента, когда система готова к повторению процесса называется рабочим циклом поперечных мостиков.

Рис. 11. Цикл взаимодействия тонких и толстых нитей: А- состояние покоя; Б- прикрепление головки миозина к актину, поворот головки миозина; В- высвобождение АДФ и Фн, развитие тянущего усилия, присоединение новой молекулы АТФ, что приводит к отделению головки миозина от молекулы актина; Г-гидролиз АТФ возвращает молекулу миозина в исходное положение.
Каждый цикл состоит из 4 стадий:
Прикрепление поперечного мостика к актиновому филаменту (образование актомиозиновых мостиков увеличивает АТФазную активность головок миозиновых нитей и освобождение энергии АТФ)
Гребковое движение (деформация актомиозиновых мостиков в шарнирных участках, что вызывает смещение актиновой нити вдоль миозиновой на 1 % длины)
Отсоединение поперечных мостиков
Получение энергии (при присоединении АТФ к головке миозина и ее гидролизе) - головка присоединяется в новом месте
Таким образом, в цикле поперечных мостиков АТФ выполняет две разные роли: 1) его гидролиз поставляет энергию для движения поперечного мостика; 2) его связывание (но не гидролиз) с миозином сопровождается отделением последнего от актина и создает возможность повторения цикла поперечных мостиков.
Расслабление мышц. В настоящее время расслабление считается активным процессом, причем расход энергии больше, чем при сокращении. Источник сокращения - АТФ, расщепляющийся при сокращении.
Доказательства:
-появление мышечных контрактур при утомлении (т. е. при деятельном сокращении);
-явления трупного окоченения (отсутствие энергии).
Для расслабления миоцита необходимы два главных условия:
Наличие достаточного уровня АТФ
Низкая концентрация Са 2+ - ниже до 10 -7 (создается активацией Са 2+-насоса, перемещение Са 2+- в цистерны саркоплазматического ретикулума, снижение концентрации Са 2+- в саркоплазме, восстановление блокады тропомиозином миозинсвязывающих участков актиновых нитей).