

Нейрохирургия
.pdfРис.
2.19. Фронтальный срез большого мозга на уровне сосцевидных тел. 1 - продольная щель большого мозга;
2 - свод; 3 - мозолистое тело; 4 - сосудистое сплетение бокового желудочка; 5 - лучистость мозолистого тела; 6 - прецентральная извилина; 7 - центральная борозда; 8 - постцентральная извилина; 9 - медиальное ядро таламуса; 10 - хвост хвостатого тела; 11 - задняя часть передней спайки; 12 - гиппокамп; 13 - неопределенная зона; 14 - субталамическое ядро; 15 - III желудочек; 16 - сосцевидные тела; 17 - сосцевидно-таламический пучок; 18 - основание ножки мозга; 19 - миндалевидное тело; 20 - зрительный тракт; 21 - нижний рог бокового желудочка; 22 - верхняя височная борозда; 23 - ограда; 24 - островок; 25 - латеральная борозда; 26 - покрышка; 27 - скорлупа; 28 - бледный шар; 29 - внутренняя капсула; 30 - латеральные ядра таламуса; 31 - хвостатое ядро; 32 - мозговая пластинка таламуса; 33 - передние ядра таламуса

и онтогенезе раньше, чем хвостатое ядро и скорлупа), гистологической структурой и функциями; подразделяется на наружный и внутренний сегменты.
Наибольшее число афферентных связей хвостатое ядро, скорлупа и бледный шар получают из коры, особенно из ее передних отделов, моторной и соматосенсорной зон. Вторым важнейшим источником афферентации этих образований являются пути, идущие от таламуса, в основном от неспецифических внутрипластинчатых и срединных ядер. Источником афферентных связей бледного шара является также средний мозг, прежде всего черное вещество. Особенно много волокон из черного вещества поступает в хвостатое ядро, а нигростриарный дофаминергический путь имеет большое функциональное значение. Имеются афферентные связи базальных ганглиев с миндалевидными телами, ретикулярной формацией, гиппокампом, мозжечком.
Эфферентные связи осуществляются через пути из бледного шара в таламус. Проекции хвостатого ядра и скорлупы на таламус в основном не прямые, а опосредованные через бледный шар. Известны также восходящие проекции от паллидума к коре, главным образом к лобным долям ипсилатерального полушария. При этом установлено перекрытие проекций волокон из хвостатого ядра, скорлупы и бледного шара в ряде полей коры большого мозга.
Существуют и каудато-паллидарные связи, а также проекции стриатума и паллидума на черное вещество, ретикулярную формацию и дру-
Рис. 2.20. Горизонтальный срез большого мозга на уровне мозолистого тела. 1 - колено мозолистого тела; 2 - свод; 3 - наружная капсула; 4 - самая наружная капсула; 5 - ограда; 6 - чечевицеобразное ядро; 7 - III желудочек; 8 - внутренняя капсула; 9 - сосудистое сплетение бокового желудочка; 10 - задняя таламическая лучистость; 11 - шпорная борозда; 12 - продольная щель большого мозга; 13 -валик мозолистого тела; 14 - задний рог бокового желудочка; 15 - латеральные ядра таламуса; 16 - медиальные ядра таламуса; 17 - передние ядра таламуса; 18 - островок; 19 - внутренняя капсула (передняя ножка); 20 - головка хвостатого ядра; 21 - передний рог бокового желудочка гие системы мозга. Сложностью организации и богатством афферентных и
эфферентных связей определяется полифункциональность структур мозга, входящих в стриопаллидарную систему. Все эти структуры играют важную роль в контроле двигательных реакций, реализации условно-рефлекторной деятельности и возникновении сложных форм поведенческих реакций.
Ограда (claustrum) представляет собой узкую пластинку серого вещества, которая располагается латеральнее чечевицеобразного ядра и отделена от него наружной капсулой.
Миндалевидное тело (corpus amigdaloideum) располагается в передневерхней части парагиппокампальной извилины и в этой зоне соприкасается с древней корой. Миндалевидное тело содержит две группы ядер: одна из них состоит из базального
илатерального ядер, другая - из кортикального, медиального, центрального ядер и ядра латерального обонятельного тракта. Эти ядра различаются по гистологическим и цитоархитектоническим особенностям. Миндалевидное тело принимает участие в корригирующем влиянии на деятельность стволовых образований, оно связано с широким диапазоном поведенческих, эмоциональных, половых, эндокринных, обменных реакций.
Внутренняя капсула - плотный слой проекционных (афферентных и эфферентных) волокон, связывающих кору большого мозга с нижележащими отделами ЦНС, ограниченный хвостатым ядром и таламусом с одной стороны, и лентикулярным ядром - с другой. На горизонтальном срезе внутренняя капсула выглядит как тупой угол, открытый кнаружи (см. рис. 2.21). Во внутренней капсуле выделяют переднюю
изаднюю ножки и соединяющее их колено. В передней ножке проходят эфферентные волокна, исходящие из коры лобной доли и направляющиеся к
таламусу (корково-таламический путь) и через мост к мозжечку (корковомостомозжечковый путь). Через колено и передние 2/3 задней ножки идут нисходящие пирамидные волокна от двигательной коры (в колене - к двигательным
ядрам черепных нервов, корково-ядерная часть пирамидного тракта; в передних 2/3 заднего бедра - к передним рогам спинного мозга, корково-
спинномозговая часть пирамидного тракта). В задней трети задней ножки проходят восходящие чувствительные волокна от таламуса к задней центральной извилине (таламокорковый путь), восходящие пути зрительного и слухового анализаторов, направляющиеся соответственно к затылочной и височной долям, а также нисходящие эфферентные волокна, идущие от нижних отделов височной и затылочной долей через мост к мозжечку (затылочно-височно-мостомозжечковый путь).
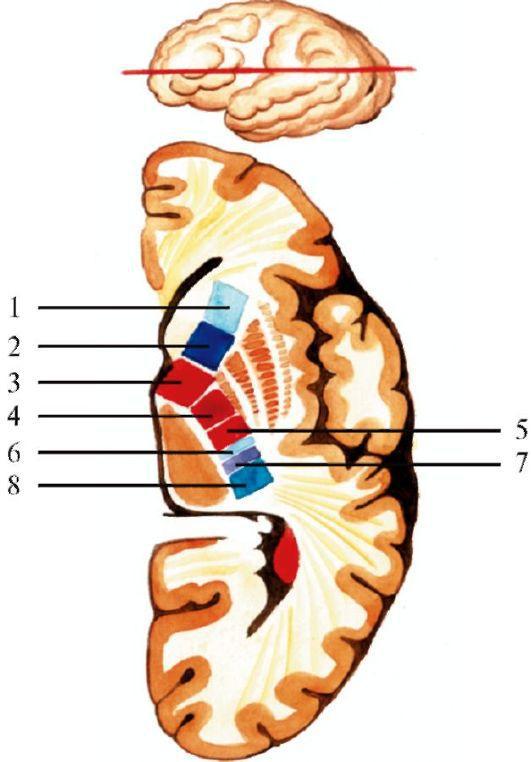
Рис.
2.21. Строение внутренней капсулы.
1 - корково-таламический путь, 2 - корково-мосто-мозжечковый путь, 3 - корковоядерная часть пирамидного тракта, 4 - корково-спинномозговая часть пирамидного тракта, 5 - таламокорковый путь, 6 - путь зрительного анализатора, 7 - путь слухового анализатора, 8 - затылочно-височно-мостомозжечковый путь Кора большого мозга
Полушария большого мозга (рис. 2.22) разделены глубокой бороздой, которая доходит до мозолистого тела - массивного слоя волокон, соединяющих оба полушария. Каждое полушарие имеет три полюса: лобный, затылочный и височный. Макроскопически в каждом полушарии выделяются лобная, теменная, затылочная, височная доли и островок.
Поверхность большого мозга образована корой, состоящей из нервных клеток. Под корой большого мозга располагается слой волокон, связывающих кору большого мозга с подкорковыми образованиями и отдельные ее поля между собой. Поверхность полушарий большого мозга изрезана бороздами, разделяющими ее на доли и извилины. Различают первичные борозды, встречающиеся постоянно, глубокие, рано появляющиеся на мозге эмбриона; вторичные, тоже постоянные, но появляющиеся позднее, и третичные, непостоянные борозды.
Среди наиболее крупных первичных борозд выделяют:
1)центральную (роландову) борозду (sulcus centralis), которая отделяет лобную долю от теменной;
2)латеральную (сильвиеву) борозду (sulcus lateralis), которая отделяет лобную и теменную доли от височной;
3)теменно-затылочную борозду (sulcus parietooccipitalis), отделяющую теменную долю от затылочной. Первые две борозды располагаются на наружной поверхности полушарий, третья - на медиальной.
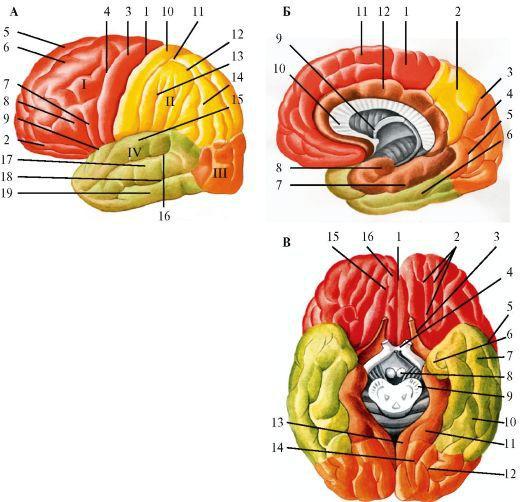
Рис.
2.22. Полушария большого мозга. А - верхнелатеральная поверхность левого полушария: 1 - центральная борозда; 2 - глазничная часть нижней лобной извилины; I - лобная доля: 3 - прецентральная извилина; 4 - прецентральная борозда; 5 - верхняя лобная извилина; 6 - средняя лобная извилина; 7 - покрышечная часть нижней лобной извилины; 8 - треугольная часть нижней
лобной извилины; 9 - латеральная борозда; II - теменная доля: 10 - постцентральная извилина; 11 - постцентральная борозда; 12 - внутритеменная борозда; 13 - надкраевая извилина; 14 - угловая извилина; III - затылочная доля; IVвисочная доля: 15 - верхняя височная извилина; 16 - верхняя височная борозда; 17 -
средняя височная извилина; 18 - средняя височная борозда; 19 - нижняя височная извилина.
Б - медиальная поверхность правого полушария: 1 - парацентральная долька; 2 - предклинье; 3 - теменно-затылочная борозда; 4 - клин; 5 - язычная извилина; 6 - латеральная затылочно-височная извилина; 7 - парагиппокампальная извилина; 8 - крючок; 9 - свод; 10 - мозолистое тело; 11 - верхняя лобная извилина; 12 - поясная извилина.
В - нижняя поверхность большого мозга: 1 - продольная щель большого мозга; 2 - глазничные борозды; 3 - обонятельный нерв; 4 - зрительный перекрест; 5 - средняя височная борозда; 6 - крючок; 7 - нижняя височная извилина; 8 - сосцевидное тело; 9 - основание ножки мозга; 10 - латеральная затылочно-височная извилина; 11 - парагиппокампальная извилина; 12 - коллатеральная борозда; 13 - поясная извилина; 14 - язычная извилина; 15 - обонятельная борозда; 16 - прямая извилина
Цитоархитектоника коры большого мозга Кора большого мозга разделяется на новую, древнюю, старую и промежуточную, существенно различающиеся по строению.
Новая кора (neocortex) занимает около 96% всей поверхности полушарий большого мозга и включает затылочную, нижнюю теменную, верхнюю теменную, постцентральную, прецентральную, лобную, височную, островковую и лимбическую области.
Новая кора большого мозга имеет шестислойное строение (рис. 2.23): слой I - молекулярная пластинка (lamina molecularis); слой II - наружная зернистая пластинка (lamina granulans externa); слой III - наружная пирамидная пластинка (lamina pyramidalis externa); слой IV - внутренняя зернистая пластинка (lamina granulans interna); слой V - внутренняя пирамидная
пластинка (laminapyramidalis interna); слой VI - мультиформная пластинка(lamina multiformis). Все слои представлены в гомотипической коре. В некоторых полях число слоев уменьшается вследствие исчезновения того или иного слоя или увеличивается в результате разделения слоя на подслои (гетеротипическая кора). Цитоархитектонические особенности различных участков коры большого мозга обусловлены шириной поперечника коры и ее отдельных слоев, величиной клеток, плотностью их расположения в различных слоях, выраженностью горизонтальной и вертикальной исчерченности, разделением отдельных слоев на подслои, другими специфическими признаками строения данного поля и т.д. Эти особенности лежат в основе разделения коры большого мозга на области, подобласти, поля и подполя. Важным критерием для классификации областей и полей коры большого мозга являются закономерности их развития в онто- и филогенезе. Эволюционный подход, который был успешно использован для изуче-

Рис.
2.23. Клеточное строение коры большого мозга.
1 - I слой; 2 - II слой; 3 - III слой; 4 - IV слой; 5 - V слой; 6 - VI слой
ния цитоархитектоники коры больших полушарий, позволил создать современную классификацию полей коры большого мозга (рис. 2.24).
Затылочная область связана с функцией зрения, имеет высокую плотность клеток, просветленный слой V, нечеткое разделение слоев II и III. В нижней теменной области большая ширина коры, высокая плотность клеток, выражены слои II и IV, радиальная исчерченность проходит через все слои. Эта область связана с наиболее сложными ассоциативными, интегративными и аналитическими функциями.
Верхняя теменная область также участвует в сложных интегративных и ассоциативных функциях, у нее горизонтальная исчерченность, средняя ширина коры, крупные клетки в слоях III и V, хорошо различимы слои II и IV. Постцентральная область связана с чувствительностью, восприятие раздражений с различных участков тела организовано соматотопически. Эта область имеет
небольшую ширину коры, выраженные слои II и IV, множество клеток во всех слоях, светлый слой V.
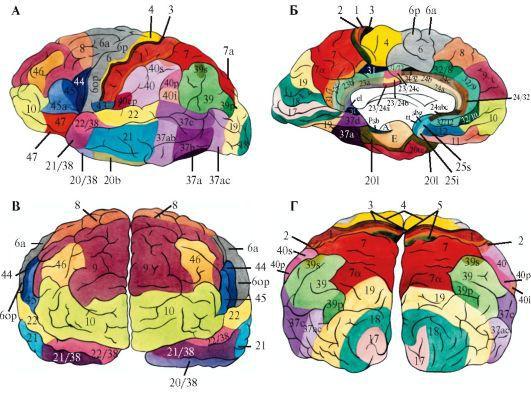
Рис.
2.24. Карта цитоархитектонических полей коры большого мозга человека (Институт мозга).
А - наружная поверхность. Б - внутренняя. В - передняя. Г - задняя. Цифрами обозначены поля
В прецентральной области слабо выражен слой II, слой IV отсутствует, в V слое очень крупные пирамидные клетки, ширина коры сравнительно большая. Эта область определяет произвольные движения; регуляция функций определенных мышц также организована соматотопически. Лобная область связана с ассоциативными и интегративными функциями, играет важную роль в высшей нервной деятельности. В этой области широкая кора, выражены слои II и IV, широкие слои III и V разделены на подслои.
Височная область связана со слуховым анализатором. Она разделяется на 4 подобласти, каждая из которых имеет особенности архитектоники. Островковая область обеспечивает функции речи, частично связана с анализом обонятельных и вкусовых ощущений. В этой области сравнительно большая ширина коры, широкий слой IV, выраженная горизонтальная исчерченность.
Лимбическая кора связана с вегетативными функциями, регуляцией эмоций. Ее цитоархитектоническая характеристика представляется весьма сложной, типичных признаков всех ее полей нет.
Древняя кора (paleocortex) включает обонятельный бугорок, диагональную область, прозрачную перегородку, периамигдалярную и препириформную области. Четкой границы с подлежащими подкорковыми образованиями нет.
Старая кора (archicortex) включает аммонов рог, зубчатую фасцию, основание аммонова рога (subiculum) и taenia tecta. Старая кора в отличие от древней коры четко отделена от подкорковых образований.
Как старая, так и древняя кора не имеет шестислойного строения. Она представлена трехслойными или однослойными структурами. Между древней, старой корой и окружающими их формациями располагаются промежуточные зоны - перипалеокортикальные формации.
2.3. Оболочки головного и спинного мозга
Выделяют твердую (dura mater), паутинную (arachnoidea) и мягкую, или сосудистую (pia mater), мозговые оболочки (рис. 2.25).
Твердая мозговая оболочка окружает мозг снаружи. Она образует ряд отростков, вдающихся между отдельными частями мозга: большой серповидный отросток (между полушариями большого мозга), малый серповидный отросток (между полушариями мозжечка), намет мозжечка (между затылочными долями и мозжечком) и диафрагму турецкого седла (см. рис. 2.24).
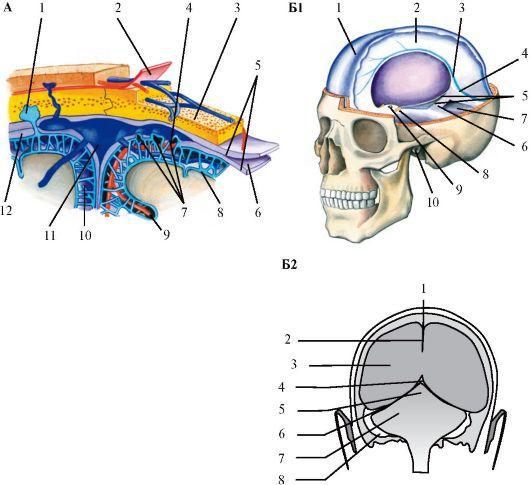 Рис.
Рис.
2.25. Оболочки головного мозга. А - Строение оболочек и подоболочечных пространств: 1 - пахионовы тельца; 2 - апоневротический шлем; 3 - диплоэ; 4 - церебральные артерии; 5 - твердая мозговая оболочка; 6 - эпидуральное пространство; 7 - паутинная оболочка; 8 - мягкая мозговая оболочка; 9 - пространство Вирхова-Робина;
10- субарахноидальное пространство;
11- верхний сагиттальный синус; 12 - субдуральное пространство. Б - Внутричерепные полости (синусы): Б1 - вид сбоку, 1 - верхний сагиттальный синус; 2 - серп большого мозга; 3 - нижний сагиттальный синус; 4 - прямой синус; 5 - края намета мозжечка; 6 - намет
мозжечка; 7 - субтенториальное пространство; 8 - диафрагма турецкрго седла; 9 - воронка; 10 - внутренний слуховой проход; Б2 - вид сзади, 1 - верхний сагиттальный синус; 2 - серп большого мозга; 3 -
супратенторальное пространство; 4 - прямой синус; 5 - серп большого мозга; 6 - намет мозжечка; 7 - субтенториальное пространство; 8 - сигмовидный синус В дупликатурах твердой мозговой оболочки располагаются венозные синусы, куда собирается венозная кровь. Синусы не имеют клапанов, поэтому возможен
обратный ток крови. К наиболее крупным венозным синусам относятся следующие
(см. рис. 2.24).
Верхний сагиттальный синус (sinus sagittalis superior) находится в верхнем крае большого серповидного отростка. Нижний сагиттальный синус
(sinus sagittalis inferior) расположен вдоль нижнего края большого серповидного отростка твердой мозговой оболочки и вливается в прямой синус (sinus
rectus). Прямой синус лежит в дупликатуре намета мозжечка, направляется спереди назад и вниз, соединяется с верхним сагиттальным синусом и вливается в поперечный синус. Поперечный синус (sinus transversus) - парный и самый крупный из всех синусов, расположен по заднему краю мозжечкового намета. У пирамид височной кости синус делает изгиб и идет дальше под названием сигмовидного синуса, который вливается во внутреннюю яремную вену.
Между мягкой мозговой оболочкой и паутинной оболочкой расположено субарахноидальное пространство, представляющее собой щелевидную полость, заполненную цереброспинальной жидкостью и содержащее кровеносные сосуды и многочисленные трабекулы. Ограничивающие пространство оболочки и содержащиеся в нем структуры выстланы плоскими клетками арахноидэндотелия
(см. рис. 2.25).
2.4. Цереброспинальная жидкость и ликвороциркуляция ЦСЖ представляет собой ультрафильтрат плазмы крови, вырабатываемый
сосудистыми сплетениями желудочков, в основном боковых (рис. 2.26). Мозг окружен ЦСЖ, которая заполняет вентрикулярную систему, цистерны, располагающиеся на основании мозга и по ходу крупных сосудов, и субарахноидальное пространство (рис. 2.27). Ее отток из вентрикулярной системы осуществляется через отверстия, соединяющие боковые желудочки с III желудочком (отверстие Монро), далее по водопроводу мозга из IV желудочка ЦСЖ поступает через срединную апертуру (отверстие Мажанди) в затылочную цистерну и через латеральную апертуру (отверстие Люшка) в боковых выворотах ГУ желудочка в цистерны моста. Резорбция ЦСЖ происходит на поверхности мозга вблизи сагиттального синуса через арахноидальные (пахионовы) грануляции. Ежедневно вырабатывается 500-750 мл ЦСЖ и такое же количество ее всасывается, в связи с чем общее количество жидкости, находящейся в черепе, остается практически неизменным (100-150 мл). При такой скорости обновление ЦСЖ происходит приблизительно 4 раза в сутки. Общий объем ЦСЖ в желудочках мозга - примерно 35 мл, а подпаутинное пространство содержит около 100 мл.