

2.2. Элемент Нернста
Природа биоэлектрогенеза была понятна лишь после создания теории электрической диссоциации (С.Аррениус 1887г.). Опыт многих исследователей убеждает, что в живых тканях, которые могут рассматриваться как растворы электролитов, генерация потенциала связана с неравномерным распределением ионов.
Самой простой и адекватной моделью является элемент Нернста (Рис.12.). Сосуд, в котором находится растворы одной и той же соли, но разной концентрации, разделен пористой перегородкой (C1 > C2). Вследствие разницы концентрации, ионы диффундируют, причем один из ионов, как правило, катион диффундирует быстрее и более разбавленный раствор приобретает знак катиона. По обе стороны перегородки образуется разность потенциалов, которую называют диффузионной разностью потенциалов.
При этом ЭДС, возникающая в концентрационном элементе, образованном раствором одной соли, определяется из уравнения Нернста.
![]()
![]() - газовая постоянная
(R=8,316 Дж/к-моль),
- газовая постоянная
(R=8,316 Дж/к-моль),
![]() - абсолютная температура,
- абсолютная температура,
![]() - число Фарадея
(96500 кл/моль), n-
валентность, С1
и С2
молекулярные концентрации
ионов по обе стороны мембраны.
- число Фарадея
(96500 кл/моль), n-
валентность, С1
и С2
молекулярные концентрации
ионов по обе стороны мембраны.
В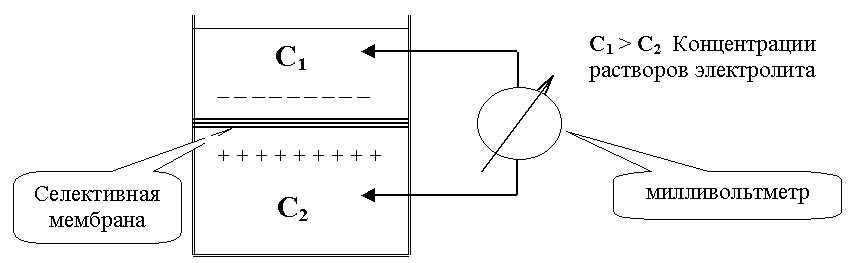 реальных условиях, в клетке, разница
скорости движения анионов и катионов
обусловлена присутствием полупроницаемой
мембраны. Без нее ЭДС концентрированного
элемента быстро падает. Поэтому
потенциалы, возникающие в таком
элементе, называются не диффузионными,
а мембранными.
реальных условиях, в клетке, разница
скорости движения анионов и катионов
обусловлена присутствием полупроницаемой
мембраны. Без нее ЭДС концентрированного
элемента быстро падает. Поэтому
потенциалы, возникающие в таком
элементе, называются не диффузионными,
а мембранными.
Рис. 12.
Элемент Нернста.
Теория электролитической диссоциации была впервые применена и объяснена в механизме биоэлектрогенеза В.Ю.Чаговцем (1896) слушателям ВМА, в дальнейшем развил учение Ю.Бернштейн и лауреаты Нобелевской премии (Ходжкин, Кац, Хакси).
2.3. Потенциал покоя.
Установлено, что цитоплазма клеток существенно отличается по химическому составу от внеклеточной жидкости, находящейся в межклеточном пространстве (рис. 13.)
-
Ионы
Среда
Внеклеточная
Внутриклеточная
K+
1
41
Na+
10
1
Cl-
13
1
Рис. 13. Относительная концентрация ионов внутри и вне клетки.
Разность ионных концентраций создает силу, стремящуюся выровнять содержание ионов, но мембрана препятствует этому процессу. Мембраны обеспечивают избирательную проницаемость для различных ионов. Вследствие этого в тканях наблюдается неодинаковая скорость диффузии через клеточные мембраны катионов и анионов, что при наличии концентрационных градиентов служит непосредственной причиной возникновения мембранных потенциалов. Их называют биопотенциалами.
Таким образом, источником электричества в живой ткани служит концентрационный элемент, образованный растворами электролита, имеющего неодинаковую концентрацию в цитоплазме и межклеточной жидкостью. При рассмотрении живых тканей в качестве концентрационного элемента необходимо учитывать диффузию через клеточную мембрану не одного иона, а всех, концентрации которых неодинаковы внутри и вне клетки. Это обстоятельство учитывает уравнение Гольдмана-Ходжкина-Катца.
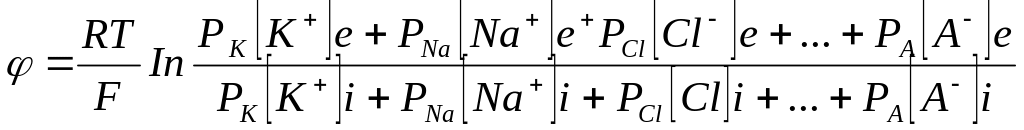 (3.3)
(3.3)
в
числителе выражения, стоящего под знаком
логарифма, представлены концентрации
![]() ,
,
![]() ,
но
,
но
![]() ,
а в знаменателе -
,
а в знаменателе -
![]() ,
,
![]() ,
но
,
но
![]() ,
так как ионы хлора отрицательно заряжены.
,
так как ионы хлора отрицательно заряжены.
![]() - проницаемость мембраны, зависящая от
определенных условий.
- проницаемость мембраны, зависящая от
определенных условий.
Уравнение Гольдмана позволяет рассчитать разность потенциалов, существующую в разных клетках между цитоплазмой и межклеточной средой в покое и при возбуждении.
Потенциал покоя - стационарная разность электрических потенциалов, регистрируемая между внутренней и наружной поверхностями мембраны в невозбужденном состоянии.
Потенциал покоя определяется разной концентрацией ионов по разные стороны мембраны и диффузией ионов через мембрану. Результаты расчетов приблизительно совпадают с величинами мембранных потенциалов, полученными опытным путем. Проанализируем уравнение Гольдмана в состояние покоя:
В состоянии покоя
проницаемость мембраны для ионов
![]() значительно больше, чем для
значительно больше, чем для
![]() ,
и больше, чем для
,
и больше, чем для
![]() :
:
![]()
Для аксона кальмара, например,
![]() Из уравнения
Гольдмана получим уравнение Нернста
для мембранного потенциала покоя:
Из уравнения
Гольдмана получим уравнение Нернста
для мембранного потенциала покоя:
![]() ;
;
![]()
Таким образом,
уравнение Нернста - частный случай
уравнения Гольдмана. При концентрации
в клетке 20 мМ и аксоплазме 400 мМ рассчитанное
значение
![]() равно - 95мВ. Действительно, в опытах на
аксонах регистрируют потенциалы покоя
около - 90мВ. Следовательно, в состоянии
покоя мембранный потенциал (ПП) равен
равновесному электрохимическому
потенциалу по
.
Важно понять, что ПП возникает за счет
более быстрой диффузии
по сравнению с
.
Разница между
равно - 95мВ. Действительно, в опытах на
аксонах регистрируют потенциалы покоя
около - 90мВ. Следовательно, в состоянии
покоя мембранный потенциал (ПП) равен
равновесному электрохимическому
потенциалу по
.
Важно понять, что ПП возникает за счет
более быстрой диффузии
по сравнению с
.
Разница между
![]() и
и
![]() определяет
направление катионного потока, создающего
ПП на мембране; катионный поток (ток
положительных зарядов) направлен из
клетки наружу (выходящий ток) (рис.14).
определяет
направление катионного потока, создающего
ПП на мембране; катионный поток (ток
положительных зарядов) направлен из
клетки наружу (выходящий ток) (рис.14).
Рис.14. двойной
слой зарядов на
клеточной мембране
в состоянии покоя.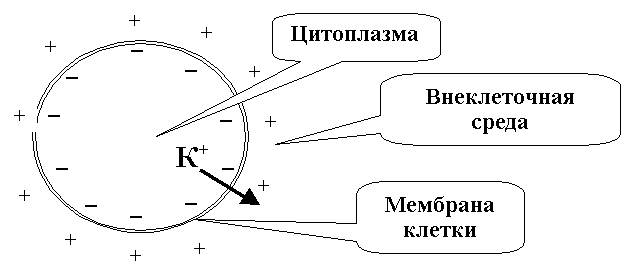 Соотношение
концентраций ионов
внутри
и снаружи клетки определяет амплитуду
ПП. Поток ионов
из клетки создает разность электрических
потенциалов между внеклеточной средой
и цитоплазмой в условиях, когда вход
в клетку и выход из нее органических
анионов (что могло бы компенсировать
нарушения электронейтральности от
потери
)
исключены свойствами покоящейся
мембраны, в этой ситуации на мембране
создается двойной электрический слой
(снаружи - катионы, главным образом
,
внутри - анионы, главным образом
органических кислот), препятствующий
дальнейшему выходу
).
Итак, все клетки в состоянии покоя
характеризуются определенной степенью
поляризации. Клеточная мембрана всегда
заряжена. ПП в различных клетках различен
и достигает нескольких десятков
милливольт. У аксона кальмара ПП
Соотношение
концентраций ионов
внутри
и снаружи клетки определяет амплитуду
ПП. Поток ионов
из клетки создает разность электрических
потенциалов между внеклеточной средой
и цитоплазмой в условиях, когда вход
в клетку и выход из нее органических
анионов (что могло бы компенсировать
нарушения электронейтральности от
потери
)
исключены свойствами покоящейся
мембраны, в этой ситуации на мембране
создается двойной электрический слой
(снаружи - катионы, главным образом
,
внутри - анионы, главным образом
органических кислот), препятствующий
дальнейшему выходу
).
Итак, все клетки в состоянии покоя
характеризуются определенной степенью
поляризации. Клеточная мембрана всегда
заряжена. ПП в различных клетках различен
и достигает нескольких десятков
милливольт. У аксона кальмара ПП![]() -85мВ,
у нервных и мышечных волокон ПП
-90мВ.
Незначительная на первый взгляд ПП
величина создает высокий градиент
потенциала (напряженность) на плазматической
мембране, т.к. толщина ее всего 10нм.
Следовательно,
-85мВ,
у нервных и мышечных волокон ПП
-90мВ.
Незначительная на первый взгляд ПП
величина создает высокий градиент
потенциала (напряженность) на плазматической
мембране, т.к. толщина ее всего 10нм.
Следовательно, ![]() это значительная напряженность
электрического поля (силовая
характеристика). Особенно важно, что
электрическое поле мембранного потенциала
покоя поддерживает закрытое состояние
так называемых активационных ворот
(см. раздел 1.4.).
это значительная напряженность
электрического поля (силовая
характеристика). Особенно важно, что
электрическое поле мембранного потенциала
покоя поддерживает закрытое состояние
так называемых активационных ворот
(см. раздел 1.4.).
Этим и обеспечивается состояние покоя и готовность клетки к возбуждению. Избирательная проницаемость плазматической мембраны всякой клетки контролирует стабильное разобщение катионов и анионов, вследствие чего на ней стойко поддерживается высокий градиент потенциала и тем самым плазматическая мембрана генерирует ЭДС. Эта ЭДС направлена против причины, ее вызывающей, и противодействует дальнейшему разобщению зарядов на мембране, т.е. выравниванию концентраций между цитоплазмой и межклеточной средой. Существование ионных градиентов на мембранах связано также с работой активного транспорта, и является энергоемким процессом.
Нарушение
биоэнергетических процессов в клетке
и работы
![]() АТФазы
приводит к уменьшению
АТФазы
приводит к уменьшению
![]() .
.
Повреждение
клеточной мембраны приводит к повышению
проницаемости клеточных мембран для
всех ионов: к повышению и
![]() и
и
![]() и
и
![]() .
Вследствие уменьшения различия
проницаемостей, абсолютное значение
мембранного потенциала
.
Вследствие уменьшения различия
проницаемостей, абсолютное значение
мембранного потенциала
![]() снижается.
снижается.
Для сильно поврежденных клеток еще меньше, но сохраняется отрицательный мембранный потенциал за счет содержащихся в клетке полианионов - отрицательно заряженных белков, нуклеиновых кислот и других крупных молекул, не могущих проникнуть через мембрану (доннановский потенциал). ПП отчасти экономит затраты свободной энергии на поддержание градиентов на биомембранах и препятствует их выравниванию. В этом одна из сторон физиологической роли ПП. Второй существенной стороной является то, что именно наличие ПП живой ткани способствует ее возбудимости!