

2.Метод падающего шарика (метод Стокса).
Метод основан на измерении скорости падения маленьких шариков в исследуемой жидкости радиусом r из вещества с плотностью в вязкой жидкости с плотностью 0 и вязкостью действуют силы:
1.сила тяжести
![]() ,
,
2.выталкивающая
сила
![]() ,
,
3.сила сопротивления жидкости, которая, согласно закону Стокса, равна FB=6 rV, где V- скорость шарика.
При равномерном движении шарика Fтяж = FA + FB , откуда
![]() gr2(ρ
- ρ0)/V
gr2(ρ
- ρ0)/V
Метод применяется при изучении оседания взвешенных частиц (крахмальных зерен, порошка какао и т. п.).
3. Ротационные методы. Измерение вязкости ротационным вискозиметром основано на определении скорости вращения цилиндра в вязкой жидкости.
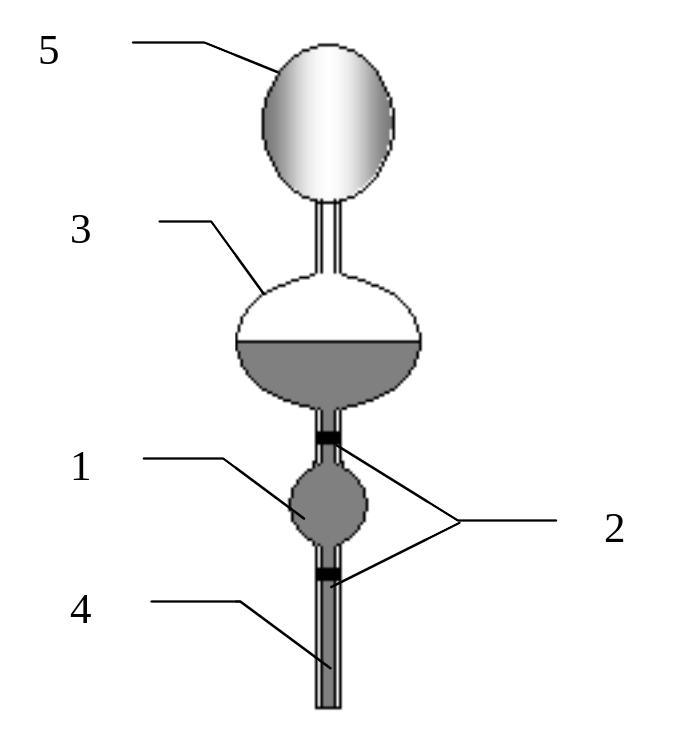
66.Устройство вискозиметра Оствальда. Определение с его помощью вязкости исследуемой жидкости.
Вискозиметр Оствальда представлен на рисунке.
С помощью вискозиметра Оствальда определяют вязкость исследуемой жидкости относительным методом. Измеряют время истечения определенного объема исследуемой и эталонной жидкостей t и t0 соответственно. Объемы жидкостей, согласно формуле, равны:

Так как перепад давления жидкости обусловлен только гидростатической силой, то р=р0=0. Выразим из формулы величину вязкости исследуемой жидкости:
![]()
1-измерительнный резервуар.2-кольцевые метки.3-резервуар.4-капилляр.5-груша.
67. Физ вопросы строения и ф-я мембран. Транспорт в-в через мембраны.
Строение и модели мембран
Все клетки окружены мембранами (цитоплазматическими, или наружными клеточными мембранами). Без мембраны содержимое клетки просто бы «растеклось», диффузия привела бы к термодинамическому равновесию, что означает отсутствие жизни. Можно сказать, что первая клетка появилась тогда, когда она смогла отделиться от окружающей среды мембраной. Внутриклеточные мембраны подразделяют клетку на ряд замкнутых отсеков (компартаментов), каждый из них выполняет определенную функцию.
Несмотря на разнообразие биологических функций и форм, все мембраны построены в основном из липидов и белков. Липидная молекула состоит из двух частей: несущей электрические заряды (полярной) головки, на которую приходится, как правило, четверть длины всей молекулы ), и длинных хвостов, не несущих электрического заряда (гидрофобных). Хвосты липидной молекулы — это длинные цепи, построенные из атомов углерода и водорода (остатки жирных кислот). Головки могут иметь разнообразное строение, однако они заряжены либо отрицательно, либо нейтральны. Связующим звеном между хвостом и головкой чаще всего служит остаток глицерина.В настоящее время наибольшее распространение имеет предложенная в 1972 г. Синджером и Николсоном жидко-мозаичная модель, в основе которой лежит все та же липидная бислойная мембрана. Эта липидная основа представляет собой как бы двумерный растворитель, в котором плавают более или менее погруженные белки. За счет этих белков полностью или частично осуществляются специфические функции мембран — проницаемость, активный перенос через мембрану, генерация электрического потенциала и т. д. Схематично жидко-мозаичная структура мембраны показана на рис. 11.3. Здесь 1 — поверхностные белки, 2 — полупогруженные белки, 3 — полностью погруженные (интегральные) белки, 4 — белки, формирующие «ионный канал» 5.
 Липиды
могут перемещаться в плоскости мембраны
(латеральная
диффузия),
а также
переходить из одного монослоя в другой
(флип-флоп
переходы). уравнение диффузии (уравнение
Фика), которое
обычно
записывают в виде:
Липиды
могут перемещаться в плоскости мембраны
(латеральная
диффузия),
а также
переходить из одного монослоя в другой
(флип-флоп
переходы). уравнение диффузии (уравнение
Фика), которое
обычно
записывают в виде:![]() D
—
коэффициент
диффузии, J-плотность
потока
D
—
коэффициент
диффузии, J-плотность
потока
68. Разновидности пассивного переноса молекул и ионов через мембраны Явления переноса относятся к пассивному транспорту: диффузия молекул и ионов в направлении их меньшей концентрации, перемещение ионов в соответствии с направлением силы, действующей на них со стороны электрического поля. Пассивный транспорт не связан с затратой химической энергии. Наиболее общая классификации видов пассивного транспорта веществ через мембрану включает в себя простую диффузию, диффузию через поры и диффузию с переносчиком. Простая диффузия через липидный бислой подчиняется уравнению Фика для молекул или, в более общем случае для нейтральных и заряженных частиц, — уравнению Нернста— Планка. В живой клетке такая диффузия обеспечивает прохождение кислорода и углекислого газа.Однако подобная простая диффузия протекает достаточно медленно и не может снабдить клетку в нужном количестве питательными веществами. Поэтому есть иные механизмы пассивного переноса веществ через мембрану, к ним относятся диффузия через канал (пору) и диффузия в комплексе с переносчиком. Два последних варианта называют иногда облегченной диффузией.Порой или каналом называют участок мембраны, включающий липидные или белковые молекулы и образующий в мембране проход .Этот канал допускает проникновение через мембрану не только малых молекул, например, молекул воды, кислорода, но и более крупных ионов. Диффузия через поры также описывается диффузионными уравнениями, однако наличие пор увеличивает коэффициент проницаемости Р.Еще одно «облегчение» диффузии — перенос ионов специальными молекулами-переносчиками .
69. Активный транспорт. Опыт Уссинга. Механизм натрий-калиевого насоса Перенос молекул в область большей концентрации, а ионов — против силы, действующей на них со стороны электрического поля пол учил название активного транспорта. Существование активного транспорта через биологические мембраны впервые было показано датским ученым Уссингом в опытах с переносом ионов натрия через кожу лягушки, которая имеет более сложную структуру, чем одиночная мембрана. Кожу лягушки можно представить как два последовательно расположенных барьера Экспериментальная камера Уссинга, разделена на две части кожей лягушки. кожа лягушки располагается между наружным и внутренним раствором: снаружи и изнутри камеры заполнены раствором Рингера, содержащим ионы натрия, калия, кальция и хлора. В результате пассивного транспорта ионы натрия диффундируют из наружного раствора в кожу. При этом цитоплазма заряжается положительно относительно этого раствора. Ионы калия, проходя из цитоплазмы во внутренний раствор, заряжают ее отрицательно. Таким образом, на коже лягушки между внутренним и внешним барьерами возникает разность потенциалов. Натрий-калиевый насос работает при условии сопряжения переноса ионов калия и натрия. Это означает, что если во внешней среде нет ионов калия, не будет активного переноса ионов натрия из клетки, и наоборот. Другими словами, ионы натрия активируют натрий-калиевый насос на внутренней поверхности клеточной мембраны, а ионы калия — на внешней. Натрий-калиевый насос переносит из клетки во внешнюю среду три иона натрия в обмен на перенос двух ионов калия внутрь клетки. Один акт переноса требует затраты энергии одной молекулы АТФ. При этом создается и поддерживается разность потенциалов на мембране, причем внутренняя часть клетки имеет отрицательный заряд.
70. Мембранные потенциалы и их ионная природа. Потенциал покоя. Мембранная теория происхождения биопотенциалов была выдвинута в 1902 г. Б. Бернштейном.Но только в 50х годах эта теория была развита и эксперементально обоснована Ходжкиным,кот принадлежат основные идеи и теории о роли ионных градиентов в возникновении биопотенциалов и о механизме распределения ионов между клеткой и средой. Сущность ее заключается в том, что потенциал, существующий на мембранах невозбужденных клеток (потенциал покоя), обусловлен полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и окружающей средой. Это распределение поддерживается механизмами активного переноса, локализованными в самой мембране.Между внутр и наружн поверхностями клеточн мембраны всегда существует разность электрич потенциалов.Эта разность потенциалов,измеренная в состояниифизиологического покоя клетки,назыв потенциалом покоя.Причиной возникновения потенциалов клеток как в покое, так и при возбуждении является неравномерное распределение ионов калия и натрия между содержимым клеток и окружающей средой. Концентрация ионов калия внутри клеток в 20 - 40 раз превышает их содержание в окружающей клетку жидкости. Напротив, концентрация натрия в межклеточной жидкости в 10 - 20 раз выше , чем внутри клеток. Такое неравномерное распределение ионов обусловлено активным переносом ионов - работой натрий-калиевого насоса.
71.Мембранные
потенциалы и их ионная природа. Уравнение
Нернста. Уравнение Гольдмана-Ходжкина-Катца.
Согласно
теории Ходжкина, Хаксли, Катца, клеточная
мембрана в состоянии покоя проницаема,
в основном, только для ионов калия.
Ионы калия диффундируют по концентрационному
градиенту через клеточную мембрану в
окружающую жидкость; анионы не могут
проникать через мембрану и остаются
на ее внутренней стороне. Так как ионы
калия имеют положительный заряд, а
анионы, остающиеся на внутренней
поверхности мембраны, - отрицательный,
то внешняя поверхность мембраны при
этом заряжается положительно, а внутренняя
- отрицательно. Понятно, что диффузия
продолжается только до того момента,
пока не установится равновесие между
силами, возникающего электрического
поля и силами диффузии. Если принять,
что потенциал покоя определяется
диффузией только ионов калия из цитоплазмы
наружу, то его величина E может быть
найдена из уравнения
Нернста:![]() ,
где [K]i и [K]e
- активность ионов калия внутри и снаружи
клетки; F - число Фародея;T - абсолютная
температура; E - изменение потенциала;
R - газовая константа.
,
где [K]i и [K]e
- активность ионов калия внутри и снаружи
клетки; F - число Фародея;T - абсолютная
температура; E - изменение потенциала;
R - газовая константа.
Если от безразмерного потенциала вернуться к электрическому потенциалу, то получаем
![]()