

7.2.3. Регенераційне відновлення
Якщо перші два шляхи післярадіаційного відновлення - репарація і репопуляція властиві всім багатоклітинним організмам, як рослинам, так і тваринам, то регенераційне відновлення - переважно лише рослинам. Еволюція прирекла більшість видів вищих рослин на прикріплений до постійного місця спосіб життя. I якщо тварини можуть укритися від несприятливих умов, які періодично виникають у навколишньому середовищі, то рослини такої можливості не мають. Саме тому поряд з такою надійною формою захисту виду, як існування надзвичайно стійкої фази їх онтогенезу – насіння, еволюція нагородила рослини потужною системою регенерації - здатності до відновлення втрачених органів і навіть всієї рослини за рахунок спеціальних тканин і органів, клітини яких, аналогічно клітинам насіння, знаходяться у стані спокою і мають високу стійкість до всіх уражуючих чинників.
Тут будуть розглянуті два основні типи регенерації у вищих рослин - регенерація тканин, які перебувають в стані спокою, і регенерація органів, які перебувають в стані спокою.
Регенерація тканин, які перебувають в стані спокою. На самому кінчику кореня, на межі з кореневим чохликом знаходиться особлива група клітин, яка має форму півсфери або двояко випуклої лінзи (рис. 7.13). Клітини цього утворення, яких нараховується не більше 1-2 тисяч, діляться дуже рідко - раз на 200-500 годин, в той час як тривалість клітинного циклу переважної кількості оточуючих їх проліферуючих клітин меристеми складає лише 12-24 години. Англійський фізіолог рослин Ф. Клаус, який відкрив і перший почав дослідження цієї ділянки кореня у 1954 році, назвав його центром спокою.
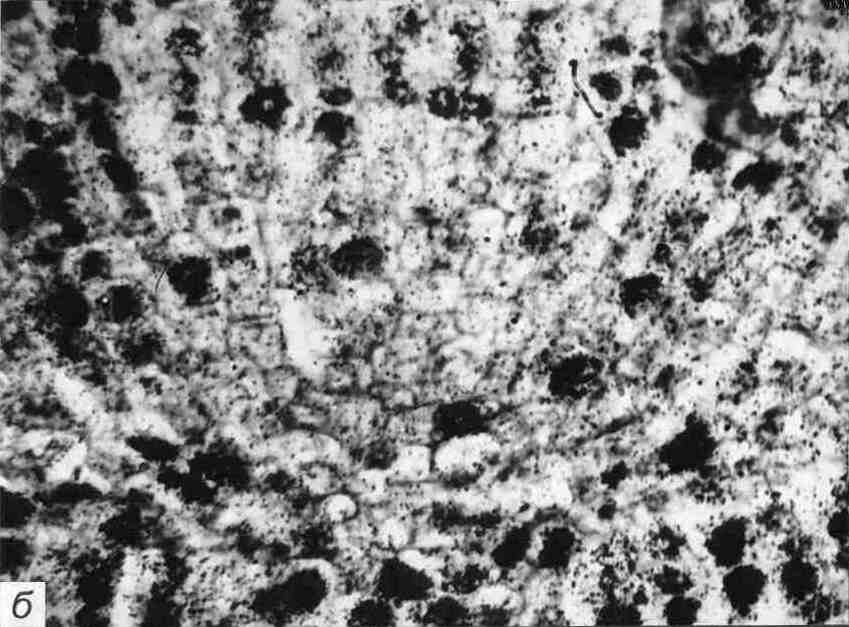

Рис. 7.13. Центр опокою у кінчику кореня: а - схематичне зображення, б - радіоавтограф зрізу кінчика кореня гороху, одержаного після витримування проростків протягом 24 год. на середовищі з 3Н-тимідіном - чітко видна зона більш світлих клітин, що не діляться, - не включають цей попередник синтезу ДНК (× 600)
Функції центру спокою до кінця не з’ясовані. Саме тому деякі ботаніки, не вбачаючи його на анатомічних препаратах за звичайних методах фарбування клітин, взагалі ігнорують його існування. Але інші дослідники вважають його основою меристеми, з якої походять всі типи клітин кореня. Деякі дослідники вважають, що клітини центру спокою є надзвичайним резервом меристеми. При пошкодженні меристеми і втраті здатності до поділу її клітин клітини центру спокою починають ділитися і відновлюють первинний клітинний об'єм.
Існує певний критичний рівень пошкодження меристеми, при якому індукується поділ клітин центру спокою. Тільки при втраті здатності до поділу у певної кількості клітин до процесів репопуляційного відновлення можуть підключатися клітини центру спокою. Очевидно, це є результатом міжклітинних взаємодій, що існують між субпопуляціями клітин, які діляться і які перебувають в стані спокою.
Після повного відновлення меристеми, коли кількість клітин, що діляться, досягає в ній рівня норми, виникає новий центр спокою.
Регенерація органів, які перебувають в стані спокою. Верхівкова меристема пагона вищих рослин, яка активно росте, в значній мірі, а іноді і повністю, придушує поділ клітин в меристемах бокових бруньок. Це явище, що отримало назву апікального домінування, являє собою сильну міжклітинну взаємодію, яка відбувається на рівні цілісного організму. Видалення верхівки приводить до зняття апікального домінування і посилення проліферативної активності тканин, які перебувають в стані спокою і пробудженню бокових бруньок. Всі ці явища належать до регенераційних.
Апікальне домінування визначається гормональним типом впливу на точки росту. Це підтверджується чисельними дослідженнями, в яких за допомогою таких рістактивуючих речовин, як кінетин і гетероауксин, вдавалося знімати апікальне домінування або, навпаки, відновлювати його при видаленій верхівковій меристемі.
Опромінення рослин іонізуючою радіацією, природно, веде до більшого пошкодження верхівкових меристем, клітини яких активно діляться і ростуть. І якщо у рослини існують органи, які перебувають в стані спокою і мають підвищену радіостійкість, то її загибель повинна викликати їх регенерацію. І, безумовно, описані в радіобіологічній літературі факти, які свідчать про сильне гілкування опромінених рослин (дихотомія, фасціації), активацію утворення бокових коренів, про підвищення кущування та інше, пояснюється саме зняттям апікального домінування і посиленням регенераційних процесів.
Найкраще апікальне домінування вивчене у бобових рослин, які здавна вважаються класичними об'єктами для його демонстрації. Бокові бруньки у бобових розміщені в пазухах листків і тому називаються пазушними. За нормальних умов росту і розвитку вони знаходяться в стані спокою протягом всього онтогенезу. Але при знятті апікального домінування, наприклад, шляхом механічного видалення верхівкової бруньки, одна (іноді дві) бокова брунька вступає в ріст і розвивається в пагін, який повністю повторює організацію головного пагона і формує в подальшому всю рослину.
Аналогічна ситуація виникає при опроміненні рослин, як, до речі, і при пошкодженні апікальної бруньки хімічними агентами, її усиханні, відмерзанні і дії інших чинників. Існує мінімальне порогове значення дози опромінення, при якій апікальне домінування повністю знімається. Безумовно, воно залежить від видової радіочутливості рослини і визначається тим рівнем дози. який веде до припинення поділу у певної критичної кількості клітин і інгібування синтезу фітогормонів, що пригнічують ріст пазушних бруньок (рис. 7.14). Саме тоді клітини меристем пазушних бруньок приступають до проліферації.

Рис. 7.14. Взаємодія фітогормонів в рослині в нормі і при знятті апікального домінування в результаті опромінення іонізуючою радіацією:
1 - рослина в нормі: цитокініни (ц), що синтезуються в корені, транспортуються в апікальну бруньку пагона, викликаючи поділ її клітин, а синтезовані в ній ауксини (а), рухаючись вниз по стеблу, за допомогою індукції утворення етилену та абсцизової кислоти пригнічують поділ клітин бокових бруньок і центру спокою кореня; 2 - при опроміненні і знятті апікального домінування пригнічується синтез ауксинів, а цитокініни спрямовуються до бокових бруньок і центру спокою, клітини яких починають поділ; 3 - в пагоні відновлюється домінування нового апікального органу, а центр спокою відновлює меристему кореня.
Регенераційне відновлення дійсно слід віднести до найпотужнішої системи відновлення, якою еволюція нагородила вищі рослини, котрі не мають можливості захищатися від пошкоджень за допомогою інших засобів. З його основі лежать неспецифічні пристосувальні реакції, які розширюють межі зміни умов життя рослин, і механізм регенераційного відновлення ефективний при дії будь-якого пошкоджуючого агента.