

Контрольные вопросы
Что такое гипогалогениты и как они образуются?
Каково строение и как функционирует миелопероксидаза?
Каково строение и биологическая роль эозинпероксидазы?
Охарактеризуйте физиологические и патологические эффекты гипогалогенитов в живых организмах?
Глава 4. Оксид азота как регулятор клеточных функций
Оксид азота (NO.) – соединение, находящееся в нормальных условиях в газообразном состоянии. Он входит в состав продуктов сгорания ископаемого топлива, тления табака, участвует в фотохимическом образовании смога. В 1986 году оксид азота был идентифицирован как эндотелиальный фактор релаксации сосудов (ЭФР), а в дальнейшем как медиатора нервной, иммунной и других систем организма, что открыло новые перспективы в понимании механизмов нормальной и патофизиологической функции данного соединения. Экспоненциальный рост публикаций по исследованию роли NO- в биологических системах (за 20-летний период публикаций свыше 40 тыс.статей) позволил Американской Ассоциации развития науки назвать в 1992 году NO- молекулой года. Открытие и изучение роли NO- как высокоэффективного регулятора в сердечно-сосудистой системе было отмечено в 1998 году Нобелевской премией в области физиологии и медицины, которой были удостоены американские исследователи Роберт Ферчготт, Ферид Мурад и Льис Игнарро.
Оксид азота – нетипичная сигнальная молекула: она газообразна, имеет неспаренный электрон на внешней разрыхляющей -орбитали, что придает NO- высокую реакционную способность. NO- легко окисляется и восстанавливается с образованием иона нитрозония (NO+) или нитроксил-аниона NO-. NO- хорошо растворим в воде. Будучи неполярным соединением, оксид азота легко диффундирует через липидный бислой мембран и мигрирует в биологических средах, является достаточно долгоживущим (среднее время жизни в биосистемах 5,6 с), после чего при участии кислорода и воды он превращается в нитриты и нитраты. Оксид азота образует стабильные комплексы с гемоглобином (нитрозил-гемоглобин) – HbNO и вступает в реакции S-нитрозилирования с сывороточным альбумином и другими SH-содержащими соединениями (глутатионом, гемоглобином) с образованием S-нитрозотиолов (RSNO, GSNO, SNOHb). В виде комплексов оксид азота транспортируется в организме и может реагировать в биосистемах с кислородом, супероксидным анион-радикалом (О2. ), ионами металлов переменной валентности.
В связи с этим функциональный ответ клетки на действие NO. Многообразен и в значительной степени зависит не только от фенотипа клетки-мишени, а также от количества NO в клетке, редокс-состояния NO. и его микроокружения. Установлено, что молекулы NO. синтезируются под действием фермента NO-синтазы (NOS) во многих клетках организма.
Оксид азота – аутокринная сигнальная молекула, так как участвует в проведении сигнала от мембранных рецепторов к молекулам внутрь клетки по гуанилатциклазному пути.
Оксид азота – паракринный эффектор, вносит вклад в согласованную работу близлежащих клеток, создавая «поле воздействия».
Оксид азота отвечает за широкий диапазон физиологических функций:
- регуляцию тонуса сосудов (эндотелиальный фактор релаксации);
- межклеточную коммуникацию;
- является нейромедиатором нового типа, участвует в модуляции нейротрансмиссии;
- участвует в регуляции иммунной цитотоксичности, осуществляя противораковое и бактерицидное действие;
- секрецию медиаторов и гормонов;
- экспрессию генов;
- регуляцию функции митохондрий;
- индукцию и ингибирование апоптоза.
Оксид азота – потенциально токсичная молекула, которая участвует во многих патологических процессах: гипертензии, сахарном диабете, сердечной недостаточности, канцерогенезе, наркомании, атеросклерозе, циррозе печени, нейродегенеративных болезнях, мужской импотенции и др.
Биосинтез оксида азота.
Оксид азота образуется в результате окисления кислородом гуанидиновой группы L-аргинина при участии фермента NO-синтазы (NOS), в реакции образуется также L-цитруллин:
NOS:NADFH, FAD
L-аргинин + О2 → L-цитруллин + NO.
FM, ВH4, гем
В процессе биосинтеза NO. участвует 5 электронов. По своему механизму реакция является монооксигеназной, в ходе которой один атом кислорода включается в молекулу субстрата (NO. ). При этом образуется промежуточный продукт N-гидрокси-L-аргинин. Для активности NOS необходимы кальций-связывающий белок кальмодулин (СаМ), а также коферменты и простетические группы: тетрагидроптерин (ВН4), НАДФН, ФАД, ФМН, гем с пентакоординированным Fe3+, Zn2+. NOS- это флавогемопротеин. В молекуле фермента выделяют 2 домена: оксигеназный (гемовый) домен на N-конце и редуктазный домен на С-конце.
Редуктазный домен включается в транспорт электронов от НАДФН:
НАДФН → ФАД → ФМН → гем (Fe3+)
Перенос электронов между ФАД и ФМН необходим для генерации стабильного флавинсемихинонного радикала и для восстановления гемопротеина (Fe3+→ Fe2+). В оксигеназном домене находится центр для связывания L-аргинина, происходит активация кислорода (О2→ О2. ) и окисление гуанидинового азота аргинина с образованием NO. и цитруллина.
Между доменами находится центр для связывания СаМ, без которого NOS неактивна и который определяет субклеточную локализацию фермента. ВН4 необходим для формирования нативной структуры фермента и для сближения кислорода, железа гема и гуанидиновой группы аргинина. NOS является гомодимером и связывание Zn2+ обязательно для образования димерной молекулы фермента. Работа фермента зависит от концентрации Са2+. Комплекс Са2+-СаМ включен в контроль переноса электронов между флавиновыми простетическими группами и гемом в оксигеназном домене.
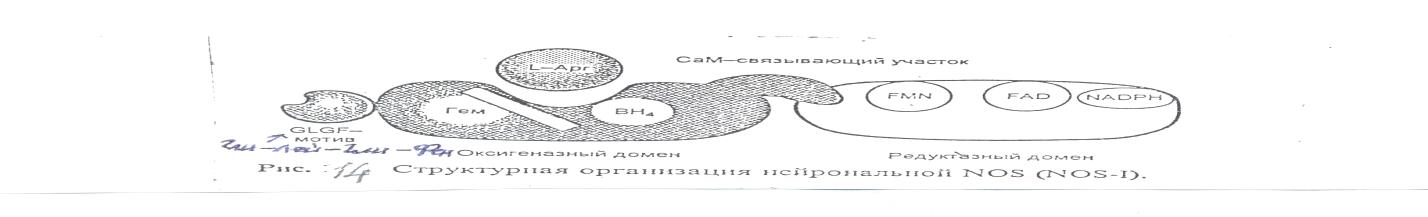
Рис. 14. Структурная организация нейрональной NO-синтазы
NOS – это семейство ферментов, образующих NO обнаружено во всех тканях и клетках. В настоящее время выделены и охарактеризованы три изоформы NOS. Две из них коститутивные (стационарные): нейрональная – nNOS (NOSI) и эндотелиальная – eNOS (NOSIII). Обнаружена еще индуцибельная форма – iNOS (NOSII).
Таблица 4
Физико-химическая характеристика NO-синтаз млекопитающих
Характеристика |
nNOS (I) |
eNOS(III) |
iNOS(II) |
Тип фермента |
Конститутивные |
Индуцибельная |
|
Источники выделения белка |
Нейроны мозга |
Эндотелий сосудов |
Макрофаги |
Уровень NO. |
Низкий – несколько мкМ |
Сотни мкМ |
|
Длительность экспрессии |
Постоянно |
Часы, дни |
|
Индукторы |
Са2+-СаМ |
Цитокинины, эндотоксины |
|
Молекулярная масса |
160 кДа |
133 кДа |
130 кДа |
Нативная структура |
Димер |
||
Локализация в клетке |
Цитозоль, митохондрии |
Цитозоль, мембрана |
Цитозоль |
Локализация в геноме |
Хромосома 12 |
Хромосома 7 |
Хромосома 17 |
Регуляция |
Обратимое фософрилирование: цАМФ-зависимые протеинкиназа (ПК), СаМ-зависимая ПК, цГМФ-зависимая ПК, ПКС |
||
Ингибиторы |
Производные аргинина |
||
В разобщенной реакции в зависимости от недостатка субстрата NOS продуцирует в дополнение к NO. активные формы кислорода – супероксид (О2. ) и перекись водорода:
НАДФН ФАД Fe3+
→ → ФАДН/ФМНН → + O2 → [Fe2+-O2] → Fe3+ + О2.
НАДФ+ ФАДН2 Fe2+
Такая активность NOS называется NOS-супероксидсинтазной, которую при недостатке аргинина в наибольшей степени проявляет nNOS. Важность этого явления заключается в том, что АФК инициируют другие сигнальные пути, отличные от NO.-зависимых, а также АФК вместе с NO. образуют новую высокоактивную частицу – пероксинитрит (ONOOH).
Таблица 5.
Некоторые ткани и клетки, экспрессирующие конститутивную и индуцибельную NO-синтазу
Конститутивная NOS (nNOS eNOS) |
Индуцибельная NOS (iNOS) |
Сосудистые эндотелиальные клетки |
Сосудистые эндотелиальные клетки |
Мезангиальные клетки |
Мезангиальные клетки |
Миокард |
Миокард |
Эндокард |
Эндокард |
Мегакариоциты |
Мегакариоциты |
Нейроны |
Остеобласты, глиальные клетки |
Периферические нервы |
Островковые клетки поджелудочной железы, сосудистые клетки гладкой мускулатуры, макрофаги (моноциты), эозинофилы, лимфоциты, фибробласты и др. |
Надпочечная железа |
|
Респираторный эпителий |
|
Тромбоциты и др. |
Многие виды клеток могут экспрессировать NOS различных типов. Получены мутантные мыши, лишенные генов каждой из NOS, но все они оказались жизнеспособными. Утрата гена NOSI привела к агрессивности и чрезмерной сексуальности; NOSII-делетированные мыши были более чувствительны к инфекциям; мыши, лишенные гена NOSIII имели повышенное давление крови. Предполагают, что функции утраченной NOS выполнялись оставшимися.
Молекулярные механизмы действия оксида азота (NO.)
Оксид азота является необычной сигнальной молекулой, так как она свободно секретируется из клетки без участия переносчиков, быстро диффундирует, не связываясь с рецепторами. Внутриклеточные эффекты NO. зависят от его редокс-состояния. С биологической точки зрения наиболее важными являются редокс-формы - NO. (2+) и NO+ (3+). Свободный радикал NO. в клетке быстро взаимодействует с молекулярным кислородом и металлами гемсодержащих и негемовых белков. Это приводит к образованию нитрозильных комплексов гемового и негемового железа, которые впервые в 1967 году были обнаружены А.Ф. Ваниным с помощью метода электронного парамагнитного резонанса.
Оксид азота после окисления или взаимодействия с металлами образует NO+, который взаимодействует с SH-группами белков. В результате в клетке при достаточном уровне тиолов под влиянием NO. происходит нитрозилирование и изменение активности металлопротеинов, а также белков, имеющих реактивные цистеины. Регуляция активности белков нитрозилированием является одним из способов контроля функции этих белков в клетке.
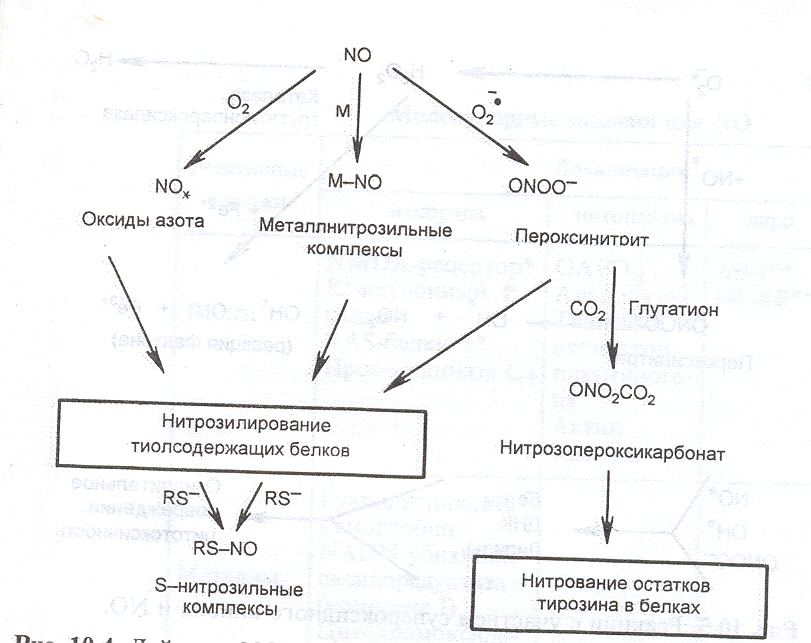
Рис. 15. Механизм действия NO. в клетке
При образовании больших количеств NO. и О2. они могут взаимодействовать с образованием пероксинитрита (ONOOH), который обладает высоким цитотоксическим потенциалом, а также способен вступать в реакцию восстановления с глутатионом и СО2, образуя нитрозопероксикарбонат (ONO2CO2), который вызывает химическую модификацию остатков тирозина в белках.
Кроме того, пероксинитрит является источником цитотоксического ОН-радикала:
О2. + NO. → ONOO- → ONOOH → OH. + NO2.
В свою очередь АКМ (ОН., NO. , ONOO-) могут окислять белки, липиды, разрушают ДНК.
Таким образом, в основе широкого разнообразия NO-эффектов в клетке лежит изменение редокс-формы NO., а также дополнительные реакции с металлами, тиолами и остатками тирозина в составе белков. Увеличение количества АФК в клетке может трансформировать эффекты NO. из защитных в цитотоксические.
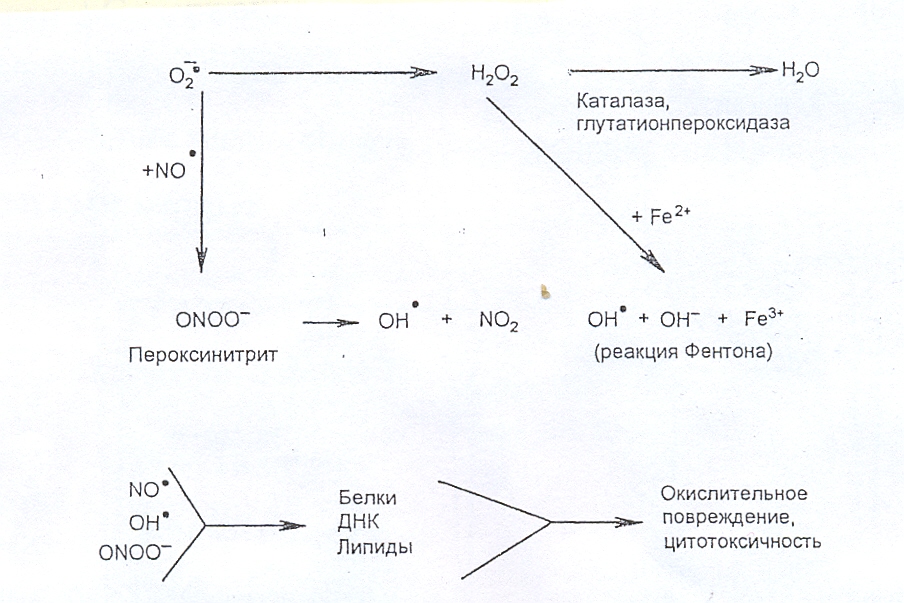
Рис. 16. Реакции с участием супероксидного аниона и NO
Молекулярные мишени для оксида азота
Реакции нитрозилирования, которые протекают путем взаимодействия активных форм кислорода с металлами, тиолами, остатками тирозина в белках изменяют их активность. Активность белков при этом может как увеличиваться (гуанилатциклаза, циклооксигеназа, транскрипционный фактор NF-KB, K+-каналы), так и уменьшаться (глицеральдегид-3-фосфатдегидрогеназа, протеинкиназа С). Белки-мишени для оксида азота разнообразны. Это мембранные рецепторы, ионные насосы, сократительные белки, ферменты и белки, участвующие в проведении сигнала.
Показано, что активация транскрипционных факторов под влиянием оксида азота увеличивает экспрессию следующих белков:
- белков теплового шока (HSP 70);
- белков антиоксидантной защиты (СОД, ГПО);
- ферритина;
- рецепторов трансферрина;
- ядерного белка р53, опухолевого супрессора;
- семейства белков Bcl-2, регуляторов апоптоза.
Таблица 6
Молекулярные мишени для NO
Реактивные участки |
Локализация |
|||
|
Мембрана |
Цитоплазма |
Ядро |
Вне клетки |
Тиолы |
NMDA-рецептор* К+-катионный канал RAS-белки*** Протеинкиназа С |
GAPD Альдолаза Тканевый Активатор Плазминогена Актин Каспазы**** |
AP-1** NF-kB** |
Глутатион Альбумин |
Металлы |
Гуанилатциклаза Гемоглобин NADH-убихинон-оксидоредуктаза (комплекс 1) Цитохромоксидаза (комплекс II) Циклооксигеназа |
|
|
|
Остатки тирозинов |
рибонуклеозилредуктаза |
|
|
|
Одной из основных белковых мишеней для оксида азота является гуанилатциклаза (ГЦ).
Гуанилатциклаза существует в двух формах: растворимой и мембраносвязанной, которые не только являются различными белками, но и отличаются механизмами регуляции. Но только растворимая ГЦ участвует в молекулярных механизмах физиологических эффектов оксида азота.
Растворимая ГЦ широко распространена и обнаружена в цитозоле практически всех клеток млекопитающих. NO. является эндогенным активатором растворимой ГЦ, что лежит в основе внутриклеточной сигнальной системы: NO.-растворимая ГЦ – цГМФ.
Растворимая ГЦ – это гем-содержащий фермент, состоящий из двух субъединиц (α и ). N-концевая часть каждой субъединицы содержит домен, связывающий гем (Fe2+). Гем отвечает за чувствительность фермента к оксиду азота. Окисление Fe2+ гема в Fe3+ снимает чувствительность к NO и может привести к потере гемовой части белка. Растворимая ГЦ – это сульфгидрильный фермент, имеющий на своей поверхности лабильные SH-группы, которые легко окисляются различными эндогенными и экзогенными окислителями и способствует активации фермента. Гем связывается с молекулой фермента через имидазольный лиганд, принадлежащий гистидину-105 -субъединицы.
Каталитический и регуляторный центры фермента разобщены и находятся в разных областях субъединиц. Каталитический участок расположен в С-концевой части α- и -субъединиц, он отвечает только за образование цГМФ и не участвует в активации фермента NO..
При активации растворимой ГЦ NO. последний взаимодействует с Fe2+ гемма с образованием комплекса нитрозил-гем. При этом атом железа выступает из плоскости порфиринового кольца и структура образовавшегося нитрозил-гемового комплекса приближается по структуре к протопорфирину IX – одному из сильных активаторов фермента (рис.16). В клетке при этом возрастает количество цГМФ, действие которого опосредуется цГМФ-зависимыми протеинкиназами и цГМФ-зависимыми фосфодитэстеразами.
Таблица 7
Механизм действия gGMP
Молекулярные мишени для gGMP |
Тип клеточного ответа |
Примеры |
Ионные каналы |
Изменение проницаемости |
Фоторецепторные клетки: открываются катионные каналы. Почки: ингибируется Na+-катионный канал |
gGMP-зависимые протеинкиназы
|
Фосфорилирование |
Медленные нейроны:увеличивается поток ионов Са2+ в нейрон. Гладкомышечные клетки: снижается [Ca2+] Тромбоциты: снижается [Ca2+] |
gGMP-стимулируемая фосфодиэстераза |
Снижение [cAMP] |
Сердце: уменьшается поток ионов Са2+ в моноцит Гипокамп: уменьшается поток ионов Са2+ (долговременное потенцирование постсинаптического ответа, формирование памяти) |
gGMP-ингибируемая фосфодиэстераза |
Увеличение [cAMP] |
Гладкомышечные клетки: снижается [Са2+] Тромбоциты: снижается [Са2+] |
Через эти ферменты цГМФ влияет на работу ионных насосов, скорость фосфорилирования белков. цГМ регулирует функцию сократительных белков, ионных насосов, регуляторных белков, участвующих в проведении сигнала, секреции медиаторов, регуляции транскрипции, трансляции и в регуляции функций митохондрий. Через цГМФ-зависимые фосфодиэстеразы регулируется концентрация цАМФ. Эти изменения проявляются в специфическом функциональном ответе на действие NO:
NO. как нейрональный мессенджер:
Роль NO. как биологического мессенджера определяется его физико-химическими свойствами. NO. является высоколабильным, короткоживущим, реактивным свободным радикалом. Долгое время считалось, что функции мозга обеспечиваются двумя типами передатчиков: возбуждающим и тормозным (ацетилхолин, катехоламины, аминокислоты, нейропептиды).
NO. является представителем нового семейства сигнальных молекул со свойствами нейротрансмиттера. Отличия NO. от традиционных нейропередатчиков:
- NO. не резервируется в синаптических везикулах нервных окончаний;
- высвобождается в синаптическую щель путем свободной диффузии, а не по механизму экзоцитоза;
- NO. синтезируется в ответ на физиологическую потребность ферментом nNOS из L-аргинина;
- биологические эффекты NO. в большой степени зависят от малой величины его молекулы, ее высокой реактивности.
Роль NO. в качестве нейромедиатора и регулятора нервной деятельности во многом связана с функциональным состоянием NМДА (N-метил-Д-аспартат)-рецепторов. Эти рецепторы опосредуют действие возбуждающих аминокислот L-глутамата и L-аспартата как нейромедиаторов, они участвуют в реализации физиологических и патогенных эффектов NO.. NМДА-рецепторы контролируют катионные Са2+-каналы в мембарне и локализованы на постсинаптических мембранах. L-глутамат, имеющий исключительно важное значение для деятельности нервной системы, синтезируется в нервных окончаниях из глюкозы и глутаиона, откладывается в синаптических везикулах, откуда высвобождается в синаптическую щель по механизму Са2+-зависимого процесса экзоцитоза. Здесь он вступает во взаимодействие с NМДА-рецепторным комплексом постсинаптической мембраны.
В физиологических условиях NМДА-рецепторы находятся в состоянии покоя, их активация глутаматом происходит при его избыточном высвобождении, вызванном усиленной стимуляцией, ведущей к деполяризации мембраны.
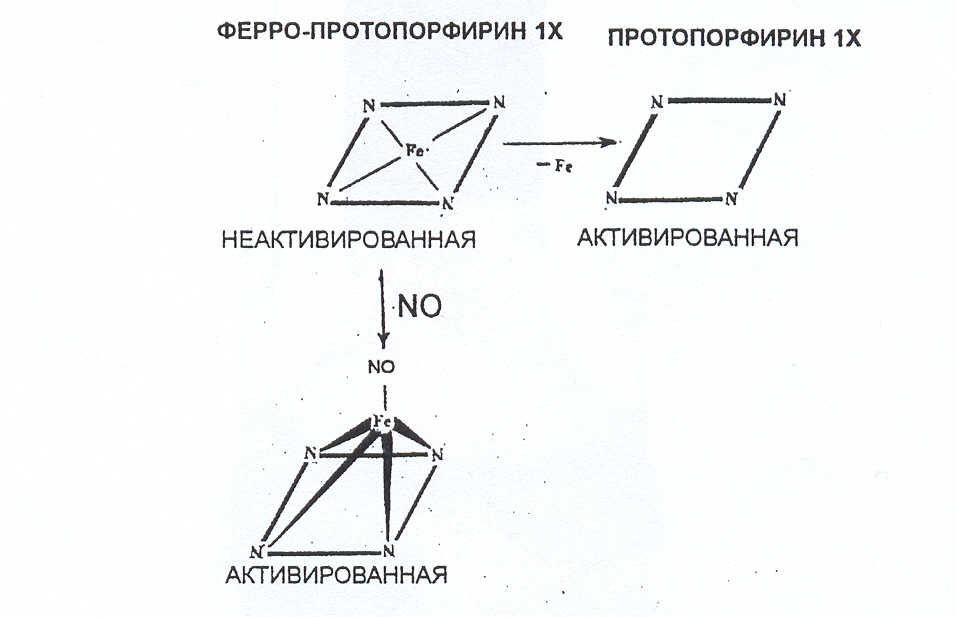
Рис. 17. Схематическое изображение активации растворимой
гуанилатциклазы NO.
В нейронах мозжечка NO. синтезируется в постсинаптических терминалах, выполняя функцию прямого нейромедиатора.
В гипокампе NO. синтезируется в постсинаптических нейронах в ответ на активацию NМДА-рецепторов. При этом вход Са2+ в клетку через Са2+-канал активирует NOS. Образующийся NO. диффундирует из постсинаптического нейрона в пресинаптический аксон, где модулирует выделение нейромедиатора глутамата. Таким образом, в гипокампе и некоторых других областях мозга NO. может выступать в роли ретроградного мессенджера. Эти механизма лежат в основе важнейшей физиологической функции NO. в головном мозгу: NO. выступает в качестве нейромедиатора как «долговременной депрессии» синапсов в мозжечке, так и феномена «длительной потенции синаптической передачи» в гипокампе, возникающих в ответ на стимуляцию рецепторов (рис.18).
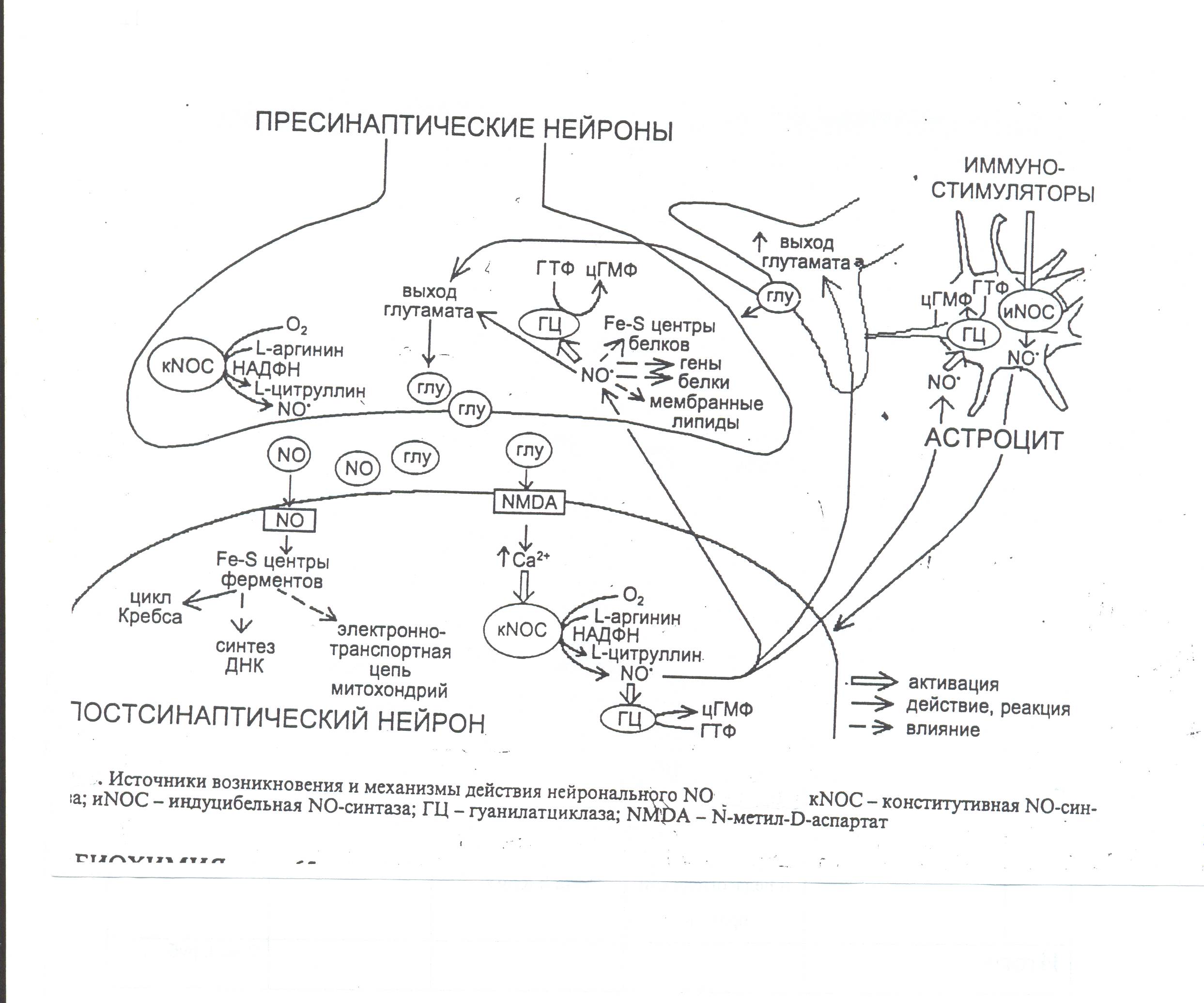
Рис. 18 Механизм нейромедиаторного действия оксида азота
NO. может принимать участие в патогенезе ряда патологий ЦНС – нейродегенеративных заболеваний, инсультов, болезни Альцгеймера и др. В этих условиях просиходит резкое увеличение высвобождения глутамата, гиперактивация ими NМДА-рецепторов и гибель окружающих участков нейронов. Непосредственной причиной гибели клеток является образующаяся в больших количествах NO., который приобретает цитотоксическое действие. Это явление получило название глутаматная нейротоксичность.
Таблица 8.