

Биосинтез днк – один из важнейших процессов передачи генетической информации последующим поколениям и ее хранения.
Некоторые их этих процессов проходят во всех типах клеток (репликация и репарация ДНК, синтез ДНК митохондрий), другие же - только в специализированных (например, осуществляющих синтез антител, транспозонов) или инфицированных вирусами клетках (репликация бактериофагов и вирусов). Все эти процессы катализируются разными ДНК- полимеразами, различающимися по числу субъединиц, молекулярной массе, ассоциацией с разными вспомогательными белками, ускоряющими процессы биосинтеза ДНК, и по многим другим структурным свойствам и функциональному назначению. В целом общие принципы синтеза ДНК схожи, однако, несомненно, существуют и различия, обусловленные видовыми и клеточными особенностями.
Как уже упоминалось выше синтез ДНК в подавляющем большинстве случаев проходит по полуконсервативному механизму, то есть дочерняя цепь ДНК синтезируется на материнской цепи, называемой обычно матрицей. Матрицами могут быть односпиральные ДНК или РНК. Ферменты, катализирующие синтез ДНК - полимеразы различаются по многим показателям. Одни из них синтезируют дочерние цепи ДНК процессивно, то есть, диссоциируя из комплекса с матричной цепью ( М-дДНК ) после удлинения дДНК на десятки нуклеотидных остатков (вплоть до полного окончания процесса), другие же функционируют дистрибютивно, диссоциируя из комплекса с матрицей практически после каждого шага удлинения. Процессивность полимераз усиливается специальными дополнительными беловыми факторами. Все ДНК-полимеразы уждаются для начала своей работы в предварительно синтезированных олигонуклеотидах – затравках, 3’-ОН группа которых используется для удлинения цепи. (Эти короткие отрезки, обычно, рибонуклеотидов синтезируются специальными РНК-полимеразами. По названию затравки – праймер эти полимеразы получили название праймаз). Все ДНК-полимеразыформируют дочерние цепи в одном направлении 5’ 3’, перемещаясь по матричной ДНК в направлении 3’ 5’. На ДНК-полимеразывозложена ответственность за точность репликации, поэтому многие из них обладают экзонуклеазной корректирующей активностью, действующей в направлении 3’ 5’. Субстратами ДНК полимераз являются дезоксирибонуклеозидтрифосфаты. Инициация репликации ДНК начинается в специальных участках со специфической нуклеотидной последовательностью (Эти участки получили название ori от origin - начало) и продолжается в двух направлениях до тех пор, пока репликативные вилки не встречаются на противоположной стороне кольцевой хромосомы бактерии. После завершения процесса две связанные кольцевые дочерние молекулы разделяются при участии специальной ферментной системы..
Скорость репликации (перемещения вилки) у бактерий составляет приблизительно 50,000-100000 пар нуклеотидов в минуту при 37 0 C. Родительская двойная спираль ДНК при такой скорости перемещения должна раскручиваться со скоростью 5,000-10000 об\мин (один виток молекулы образуется 10 парами азотистых оснований). Такой простой расчет указывает на определенные чисто механические проблемы, возникающие в процессе репликации. Основные участники процесса репликации и их основные функции у прокариот перечисляются в следующей таблице.
|
Основные ферменты и белки, участвующие в процессе репликации |
|
|
Белок |
Функция |
|
DnaB белок |
Главный белок праймосомы раскручивает ДНК в сайте ori |
|
DnaC белок |
Образует комплекс с dnaB |
|
Белок Rep |
Хеликаза (раскручивает ДНК в репликационной вилке) |
|
Праймаза (dnaG) |
Синтезирует РНК-затравку |
|
ДНК-полимеразаIII |
Полимераза, катализирующая репликацию |
|
SSB |
Связывается с одноцепочечной ДНК в репликативной вилке для предотвращения образования двойной спирали |
|
ДНК-полимеразаI |
Удаляет РНК-затравку, заполняя пробел |
|
ДНК лигаза |
Катализирует образование 3 ’, 5 -фосфодиэфирной связи, соединяя фрагменты Оказаки |
|
Топоизомераза I |
Ослабляет суперспирализацию ДНК |
|
Топоизомераза II |
Разделяет ДНК после репликации |
Последовательность событий в механизме репликации у Е Coli следует строго определенному сценарию.
oric - инициирующая последовательность E.Coli длиной в 245 пар азотистых оснований имеет участок с четырьмя повторами последовательности из 9- пар оснований и трех повторов из 13 пар оснований. Последние характеризуются высоким содержанием АТ пар и таким образом денатурируются относительно легко. К этому участку присоединяется белок - продукт гена DnaA.и специфические основные белки (HU и IHF) которые облегчают изгиб ДНК, образование которого - важный шаг в инициации репликации:
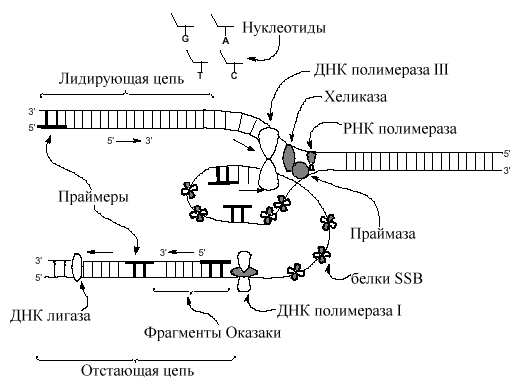
атем в дело вступает ДНК-полимераза III. Как оказалось, выше упомянутая ДНК полимераза открытая А.Корнбергом и теперь называемая ДНК-полимеразаI (Pol I) - не основной фермент репликации, хотя и играет существенную роль в процессе репликации (см. позже). У E. coli выделены две дополнительных ДНК-полимеразы, полимераза II (Pol II) и полимераза III (Pol III). Точная функция полимеразы II в клетке пока не известна. Основным ферментом, катализирующим репликацию, является полимераза III. Помимо полимеразной активности все бактериальные ДНК-полимеразыобладают 3 '-5 ' экзонуклеазной активностью. Способность катализировать гидролиз фосфодиэфирных связей и удалять нуклеотиды с 3’ концов, действуя в противоположном от своей полимеразной активности направлении, позволяет полимеразам проверять и устранять ошибки своей деятельности.
Если к растущей цепи добавляется неправильный нуклеотид, он немедленно удаляется до завершения синтеза. У полимеразы I и III обнаружена и 5 '-3 ' экзонуклеазная активность, что позволяет им участвовать в процессах репарации ДНК. Эта же экзонуклеазная активность полимеразы I используется также для удаления РНК- затравок, которые инициируют синтез цепей ДНК (см. ниже). Работе ДНК- полимеразы предшествует первичный отбор комплементарного матрице дНТФ, который проходит в общем одинаково для разных ДНК-полимераз Этот отбор начинается с образования канонических Уотсон-Криковских пар оснований. Их образование энергетически более выгодно, поскольку при этом образуется оптимальное по сравнению с другими возможными вариантами число водородных связей. Кроме того, правильно образованные пары оказывают влияние на конформационное изменение ДНК-полимеразы, направленное на ускорение реакции элонгации и последующего перемещения фермента. При образовании неправильной пары резко ухудшается кинетика процесса. Скорость перемещения фермента замедляется. При кинетической задержке элонгации цепи , возникающей в результате включения некомплементарного 3'-нуклеотидного остатка и возникновения ошибочной пары начинает проявляться активность другого центра ДНК полимеразы, расположенного вблизи центра полимеризации и, по-видимому, частично совмещающегося с ним. Этот активный центр, катализирующий 3'->5'-экзонуклеазный гидролиз удаляет некомплементарный 3'-концевой нуклеотидный остаток. Ошибочные пары возникают с частотой порядка 1: 104 - 1: 106. Дело в том, что ДНК-полимеразе на каждом шагу элонгации дДНК приходится выбирать один из четырех различающихся по структуре дНТФ в условиях, когда матричный нуклеотидный остаток в образующейся паре также вариабелен из-за соседних нуклеотидных остатков, влияющих на конформацию односпирального участка матрицы в активном центре. Редактирующая активность фермента понижает частоту ошибок в 10-100 раз.
ДНК-полимераза копирует антипараллельные цепи ДНК перемещаясь в 5 ' 3 ' направлении
Как можно объяснить, что ДНК-полимераза копирует антипараллельные цепи матрицы ДНК в одном 5 ' 3 ' направлении одновременно? Предложена модель, согласно которой ДНК-полимераза формирует димер, связанный с другими необходимыми в репликативной вилке белками (реплисома) . Одна из цепей матричной ДНК, названная отстающей цепью, временно образует петли вокруг реплисомы так, что димер ДНК-полимеразы получает возможность перемещаться по обеим цепям в одном 3' 5 ' направлении одновременно на короткое расстояние. Мономер ДНК-полимеразы, копирующий эту цепь обладает низкой процессивностью и через небольшой промежуток времени, синтезировав короткий отрезок ДНК, покидает матричную отстающую цепь. Эти короткие (до 1000 пар нуклеотидов) отрезки ДНК, образующиеся на отстающей цепи, получили название фрагментов Оказаки по имени Ф. Оказаки, впервые (1968) указавшего на прерывистый характер синтеза ДНК. Новое присоединение мономера ДНК-полимеразык матрице происходит после предварительного синтеза нового праймера. Вторая половина димера ДНК-полимеразыреплисомы , обладающая более высокой процессивностью, в чем ей помогают специфические белки, копирует дочернюю цепь ДНК без перерывов. Матричную цепь, которая копируется без перерывов, назвали ведущей цепью. Так как репликативная вилка довольно быстро (1000 нулеотидов в сек) продвигается по матрице вновь синтезируемые дочерние цепи и родительские цепи матрицы сразу формируют двойные спирали ДНК. Предполагается, что только маленький отрезок матричной двойной спирали находится в одноцепочечном состоянии в данный отрезок времени. Праймеры ведущей и отстающей цепей (10-12 пар нуклеотидов) удаляются ДНК полимеразой I, обладающей репарирующей функцией с одновременной заменой рибонуклеотидов дезоксирибонуклеотидами. Промежутки, которые возникают между 3 '-ОН и 5 '- фосфатом , воссоединяются лигазами ДНК, завершая тем самым процесс репликации.