

Защита клеток от действия протеаз
Клетки поджелудочной железы защищены от действия пищеварительных ферментов тем, что:
эти ферменты образуются в клетках поджелудочной железы в неактивной форме и активируются только после секреции в просвет кишечника.
в клетках поджелудочной железы присутствует белок-ингибитор трипсина, образующий с активной формой фермента (в случае преждевременной активации) прочный комплекс.
В полости желудка и кишечника протеазы не контактируют с белками клеток, поскольку слизистая оболочка покрыта слоем слизи, а каждая клетка содержит на наружной поверхности плазматической мембраны полисахариды, которые не расщепляются протеазами и тем самым защищают клетку от их действия.
Разрушение клеточных белков протеазами происходит при язвенной болезни желудка или двенадцатиперстной кишки.
Регуляция желудочно-кишечной секреции
Натощак секретируется незначительное количество желудочного сока.
Регуляция секреции желудочного сока осуществляется в 3 фазы:
1. Мозговая (сложнорефлекторная) фаза. Осуществляется через комплекс условных и безусловных рефлексов. Вид, запах и вкус пищи активируют нейронывагусав центре регуляции желудочной секреции. Окончания вагуса в желудке выделяютацетилхолин, который через М-холинорецепторы стимулирует синтез желудочного сока (главными, обкладочными и добавочными клетками), а также стимулирует выработку в желудке гормоновгастринаигистамина;
2. Желудочная (нейро-гуморальная) фаза. Возникает при нахождении пищи в желудке. За счетвагуса, метасимпатической нервной системы, гастрина, гистамина и питательных веществ(белки, пептиды, АК) стимулируется секреция желудочного сока. (Метасимпатическая нервная система (МНС) представляет собой комплекс микроганглиев, расположенных в стенках внутренних органов. МНС координирует и регулирует моторную, секреторную, абсорбционную, эндокринную, иммунную функции полых внутренних органов).
3. Кишечная фаза. При недостаточной обработки пищи из кишечника возникают сигналы, стимулирующие желудочную секрецию (за счет рефлексов местных и центральных, возникающих с рецепторов кишечника и реализующихся через вагус, МСН, гастрин, гистамин). При избыткеHCl или чрезмерном разрушении пищевых продуктов, из кишечникавозникают сигналы, тормозящие желудочную секрецию (через секретин, холецистокинин, ВИП, ГИП).
Гастрин – гормон пептидной природы, производимый G-клетками желудка (гастрин-17 из 17 аминокислот, и гастрин-14 из 14 аминокислот), расположенными в основном в антральном отделе желудка.
Секрецию гастрина стимулируют:
Ацетилхолин вагуса;
белки,продукты гидролиза;
бомбензин;
инсулин;
адреналин (слабо);
высокий уровень глюкокортикоидов;
гиперкальциемия.
Секрецию гастрина угнетают:
высокий уровнень HCl в желудке;
холецистокинин;
секретин;
глюкагон;
серотонин;
ГИП;
ВИП;
простагландин Е;
эндогенные опиоиды — эндорфины и энкефалины;
аденозин;
кальцитонин;
соматостатин (сильно);
Эффекты гастрина:
Гастрин связывается с гастриновыми рецепторами в желудке и активирует через аденилатциклазную систему синтез желудочного сока: он стимулирует секрецию НС1, пепсиногена, бикарбонатов и слизи в слизистой желудка.
Гастрин увеличивает продукцию простагландина E в слизистой желудка, что приводит к местному расширению сосудов, усилению кровоснабжения и физиологическому отёку слизистой желудка и к миграции лейкоцитов в слизистую. Лейкоциты принимают участие в процессах пищеварения, секретируя различные ферменты и производя фагоцитоз.
Гастрин тормозит опорожнение желудка, что обеспечивает достаточную для переваривания пищи длительность воздействия соляной кислоты и пепсина на пищевой комок.
Рецепторы к гастрину имеются и в тонкой кишке и поджелудочной железе. Гастрин увеличивает секрецию секретина, холецистокинина, соматостатина и ряда других гормонально активных кишечных и панкреатических пептидов, а также секрецию кишечных и панкреатических ферментов. Тем самым гастрин создаёт условия для осуществления следующей, кишечной, фазы пищеварения.
Гистамин биогенный амин, образующийся в энтерохромафиноподобных клетках (ECL) при декарбоксилировании аминокислоты гистидина. Секрецию гистамина стимулирует ацетилхолин вагуса, гастрин, ингибируетHCl.Гистамин, через Н2-рецепторы, усиливает секрецию HCl обкладочными клетками.
Простогландины вырабатываются покровными эпителиоцитами.Секрецию простогландинов стимулируетHCl, ингибируют глюкокортикоиды. Простогландины стимулируют слизеобразование, секрецию бикарбонатов (нейтрализация рН), усиливают кровообращения в желудке.
Серотонин– биогенный амин, образуется вэнтерохромафинных эндокриноцитах(ЕС) из 5-окситриптофана. Секрецию серотонина стимулируетHCl. Серотонин стимулирует секреторную (главные и слизистые клетки) и двигательную активность (миоциты) клеток желудка.
Соматостатин (пептид)образуется вD-клетках. Соматостатин ингибирует синтез ферментов, гормонов, соляной кислоты, увеличивает скорость всасывания воды и электролитов в тонкой кишке, снижает концентрацию вазоактивных пептидов в крови, уменьшает частоту актов дефекации и массу кала.
Пища, поступающая в желудок, стимулирует повышенное образование желудочного сока в течение 4-6 часов. Количество, состав и свойства желудочного сока меняются в зависимости от характера пищи, а также при заболеваниях желудка, кишечника и печени.Наибольшее количество желудочного сока выделяется на белковую пищу, меньше – на углеводную, еще меньше на жирную.
Регуляция поджелудочной секреции
Регуляция секреции поджелудочного сока осуществляется в 3 фазы:
1. Мозговая (сложнорефлекторная) фаза. Осуществляется через комплекс условных и безусловных рефлексов. Вид, запах и вкус пищи активируют нейронывагусав центре регуляции панкреатической секреции. Окончания вагуса в поджелудочной железе выделяютацетилхолин, который стимулирует синтез панкреатического сока.
2. Желудочная (нейро-гуморальная) фаза. Возникает при нахождении пищи в желудке. За счетвагуса, гастрина, серотонинастимулируется секреция поджелудочного сока.
3. Кишечная фаза.Кислый химус вызывает в кишечнике выделение S-клетками секретина (белковый гормон). Секретин поступают в кровь и стимулирует выделение из поджелудочной железы в тонкий кишечник панкреатического сока, содержащего много НСО3-, что нейтрализует НС1 желудочного сока и ингибирует пепсин. В результате рН возрастает от 1,5-2,0 до 7,0.
Поступление пептидов в тонкий кишечник вызывает секрецию холецистокинина (белкового гормона) в I-клетках, который стимулирует выделение панкреатического сока с большим содержанием ферментов.
Регуляция кишечной секреции
Регуляция деятельности желез тонкой кишки осуществляется местными нервно-рефлекторными механизмами, а также гуморальными влияниями и ингредиентами химуса. Механическое раздражение слизистой оболочки тонкой кишки вызывает выделение жидкого секрета с малым содержанием ферментов. Местное раздражение слизистой кишки продуктами переваривания белков, жиров, соляной кислотой, панкреатическим соком вызывает отделение кишечного сока, богатого ферментами. Усиливают кишечное сокоотделение ГИП, ВИП, мотилин. Гормоны энтерокринин и дуокринин, выделяемые слизистой оболочкой тонкой кишки, стимулируют соответственно секрецию либеркюновых и бруннеровых желез. Тормозное действие оказывает соматостатин.
Мотилин
(в Мо-клетках) - стимулирует активность
гладко-мышечной клеток кишечника.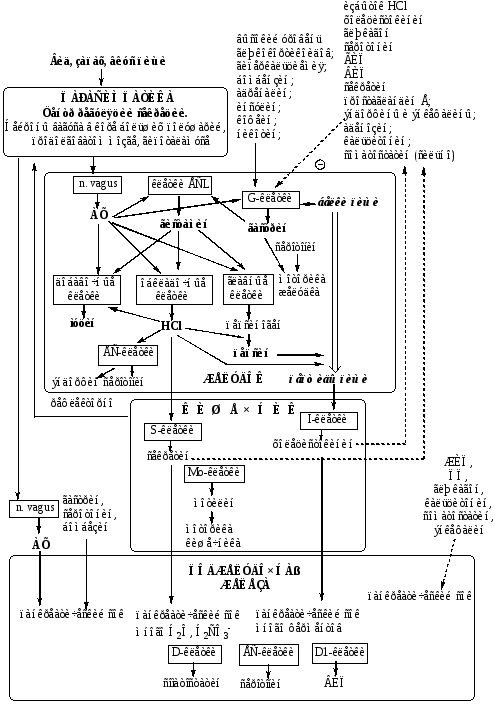
ВСАСЫВАНИЕ АМИНОКИСЛОТ В КИШЕЧНИКЕ
Всасывание L-аминокислот (но не D) — активный процесс, в результате которого аминокислоты переносятся через кишечную стенку от слизистой её поверхности в кровь.
Известно пять специфических транспортных систем, каждая из которых функционирует для переноса определённой группы близких по строению аминокислот:
нейтральных, короткой боковой цепью (аланин, серии, треонин);
нейтральных, с длинной или разветвлённой боковой цепью (валин, лейцин, изолейцин);
с катионными радикалами (лизин, аргинин);
с анионными радикалами (глутаминовая и аспарагиновая кислоты);
иминокислот (пролин, оксипролин).
Существуют 2 основных механизма переноса аминокислот: симпорт с натрием и γ-глутамильный цикл.
1. Симпорт аминокислот с Na+.
Симпортом с Nа+ переносятся аминокислоты из первой и пятой группы, а также метионин.
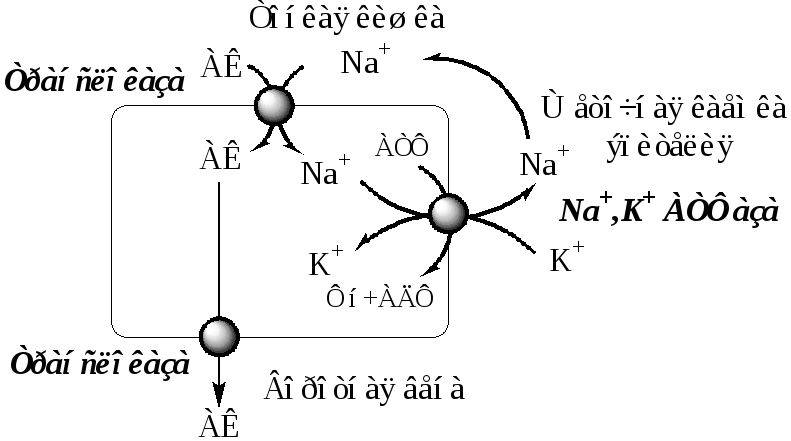
L-аминокислота поступает в энтероцит путём симпорта с ионом Na+. Далее специфическая транслоказа переносит аминокислоту через мембрану в кровь. Обмен ионов натрия между клетками осуществляется путём первично-активного транспорта с помощью Na+, К+-АТФ-азы.
2. γ-Глутамильный цикл.
γ-глутамильный цикл переносит некоторые нейтральные аминокислоты (фенилаланин, лейцин) и аминокислоты с катионными радикалами (лизин) в кишечнике, почках и, по-видимому, мозге.
В этой системе участвуют 6 ферментов, один из которых находится в клеточной мембране, а остальные — в цитозоле. Мембранно-связанный фермент γ-глутамилтрансфераза (гликопротеин) катализирует перенос γ-глутамильной группы от глутатиона на транспортируемую аминокислоту и последующий перенос комплекса в клетку. Амнокислота отщепляется от у-глутамильного остатка под действием фермента у-глутамилциклотрансферазы.
Дипептид цистеинилглицин расщепляется под действием пептидазы на 2 аминокислоты — цистеин и глицин. В результате этих 3 реакций происходит перенос одной молекулы аминокислоты в клетку (или внутриклеточную структуру). Следующие 3 реакции обеспечивают регенерацию глутатиона, благодаря чему цикл повторяется многократно. Для транспорта в клетку одной молекулы аминокислоты с участием у-глутамильного цикла затрачиваются 3 молекулы АТФ.
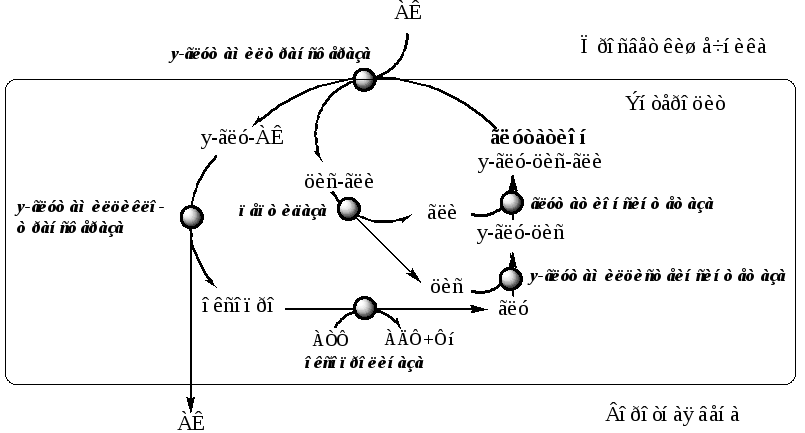
Поступление аминокислот в организм осуществляется двумя путями: через воротную систему печени, ведущую прямо в печень, и по лимфатическим сосудам, сообщающимся с кровью через грудной лимфатический проток. Максимальная концентрация аминокислот в крови достигается через 30—50 мин после приёма белковой пищи (углеводы и жиры замедляют всасывание аминокислот). Аминокислоты при всасывании конкурируют друг с другом за специфические участки связывания. Например, всасывание лейцина (если концентрация его достаточно высока) уменьшает всасывание изолейцина и валина.