

III. Обмен гормонов, производных аминокислот
К производным тирозина относятся тиреоидные гормоны и катехоламины, их обмен имеет свои специфические особенности.
Обмен катехоламинов Симпато-адреналовая ось
Подобно задней доле гипофиза, мозговой слой надпочечников — производное нервной ткани. Его можно рассматривать как продолжение симпатической нервной системы, так как преганглионарные волокна чревного нерва оканчиваются на хромаффинных клетках мозгового слоя надпочечников.
При стимуляции преганглионарного нейрона хромаффинные клетки продуцируют катехоламины — дофамин, адреналин и норадреналин.
1. Синтез. Синтез катехоламинов происходит в цитоплазме и гранулах клеток мозгового слоя надпочечников. Катехоламины сразу образуются в активной форме. Норадреналин образуется в основном в органах, иннервируемых симпатическими нервами (80% от общего количества).

2. Хранениекатехоламинов происходит в секреторных гранулах. Катехоламины поступают в гранулы путём АТФ-зависимого транспорта и хранятся в них в комплексе с АТФ в соотношении 4:1 (гормон-АТФ).
3. Секрециягормонов из гранул происходит путём экзоцитоза. Катехоламины и АТФ освобождаются из гранул в том же соотношении, в каком они сохраняются в гранулах. В отличие от симпатических нервов, клетки мозгового слоя надпочечников лишены механизма обратного захвата выделившихся катехоламинов.
4. Транспорт.В плазме крови катехоламины образуют непрочный комплекс с альбумином. Адреналин транспортируется в основном к печени и скелетным мышцам. Норадреналин лишь в незначительных количествах достигает периферических тканей.
5. Действие гормонов. Катехоламины регулируют активность ферментов, они действуют через цитоплазматические рецепторы (гликопротеины). Адреналин через α-адренергические и β-адренергические рецепторы, норадреналин – через α-адренергические рецепторы. Через β-рецепторы активируется аденилатциклазная система, через α2-рецепторы ингибируется. Через α1-рецепторы активируется инозитолтрифосфатная система. Эффекты катехоламинов многочисленны и затрагивают практически все виды обмена.
7. Инактивация. Основная часть катехоламинов быстро метаболизируется в различных тканях при участии специфических ферментов.

У катехоламинов Т½ = 10—30с. Лишь небольшая часть адреналина (~ 5%) выделяется с мочой.
Патология катехоламинов. Основная патология мозгового вещества надпочечников –феохромотитома, опухоль хромаффинных клеток. Образуется избыток катехоламинов, который проявляется повторяющимися приступами головной боли, серцебиения, потливости, повышением АД.
Обмен тиреоидных гормонов Гипоталамо-гипофизарно-тиреоидная ось
Синтез тиреоидных гормонов (йодтиронины: 3,5,3'-трийодтиронин (три-йодтиронин, Т3) и 3,5,3',5'-тетрайодтиронин (Т4, тироксин)) происходит в клетках и коллоиде щитовидной железе.
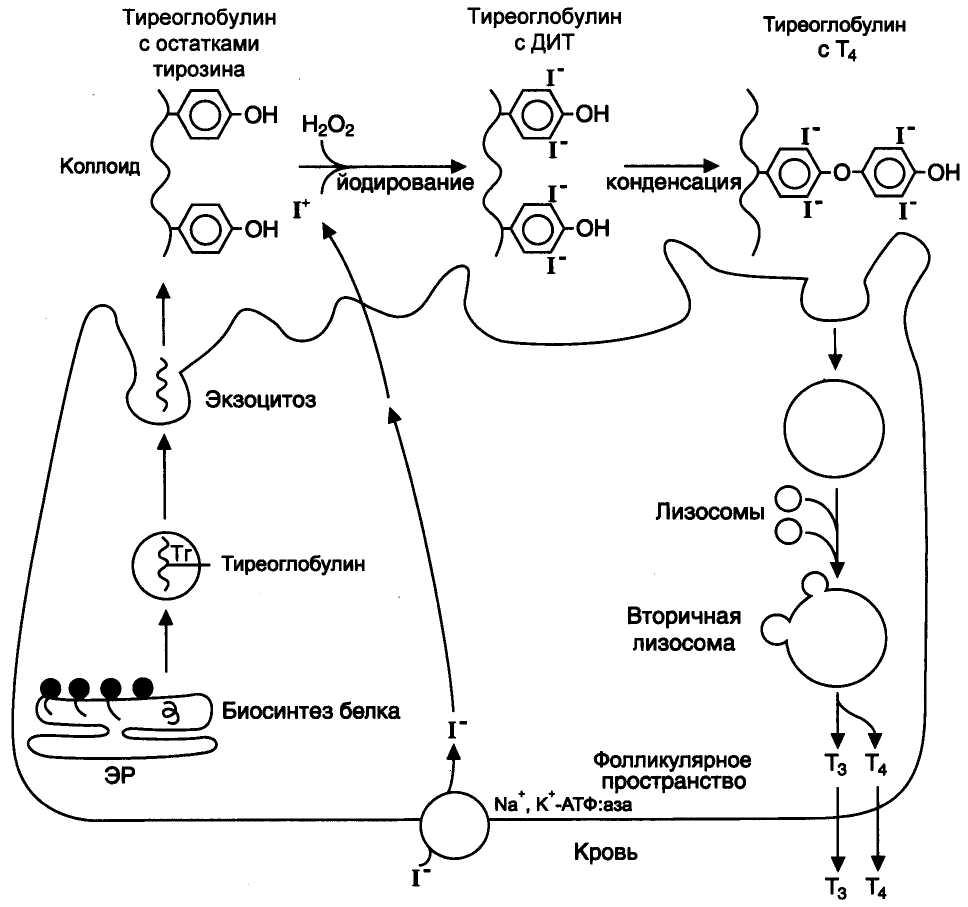
2. Транспорт йода в коллоид щитовидной железы. Йод в виде органических и неорганических соединений поступает в ЖКТ с пищей и питьевой водой. Суточная потребность в йоде 150-200 мкг. 25—30% этого количества йодидов захватывается щитовидной железой.J-поступает в клетки щитовидной железы активным транспортом при участии йодид-переносящего белка симпортом сNа+. ДалееJ-пассивно по градиенту поступает в коллоид.
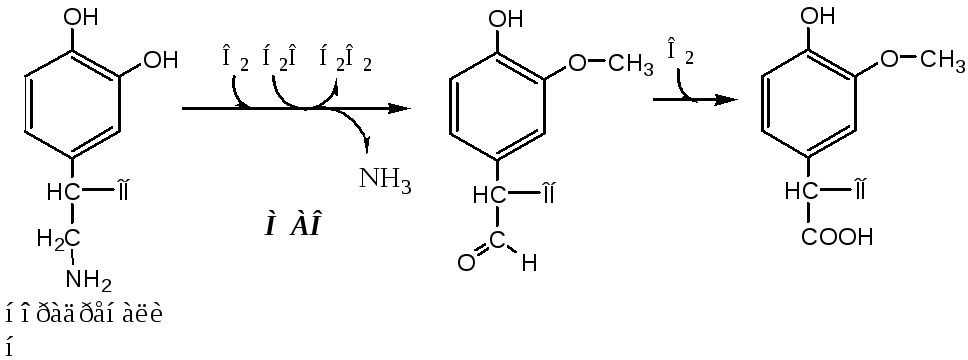
3. Окисление йода и йодирование тирозина. В коллоиде при участии гемсодержащей тиреопероксидазы и Н2О2 J-окисляется вJ+, который йодирует остатки тирозина в тиреоглобулине с образованием монойодтирозинов (МИТ) и дийодтирозинов (ДИТ).
4. Конденсация МИТ и ДИТ. Две молекулы ДИТ конденсируются с образованием йодтиронина Т4, а МИТ и ДИТ — с образованием йодтиронина Т3.
2. Хранение. В составе йодтиреоглобулина тиреоидные гормоны накапливаются и хранятся в коллоиде.
3. Секреция. Йодтиреоглобулин фагоцитируется из коллоида в фолликулярную клетку и гидролизуется в лизосомах с освобождением Т3и Т4и тирозина и других АК. Аналогично стероидным гормонам, водонерастворимые тиреоидные гормоны в цитоплазме связываются со специальные белками, которые переносят их в состав клеточной мембраны. В норме щитовидная железа секретирует 80—100 мкг Т4 и 5 мкг Т3в сутки.
4. Транспорт. Основная часть тиреидных гормонов транспортируется в крови в связанной с белками форме. Основным транспортным белком йодтиронинов, а также формой их депонирования служит тироксинсвязывающий глобулин (ТСГ). Он обладает высоким сродством к Т3и Т4и в нормальных условиях связывает почти всё количество этих гормонов. Только 0,03% Т4и 0,3% Т3находятся в крови в свободной форме.
5. Действие гормонов. Биологическая активность йодтиронинов обусловлена свободной фракцией. Основная биологически активная форма йодтиронинов - Т3; его сродство к рецепторам клеток-мишеней в 10 раз выше, чем у Т4. Дийодирование в печени Т4 до Т3по 5' увеличиваетактивностьйодтиронинов.
Йодтиронины взаимодействуют с высокоспецифичными ядерными рецепторами и регулируют экспрессию генов.
Йодтиронины участвуют в регуляции многих процессов метаболизма, развития и клеточной дифференцировки.
При физиологической концентрации йодтиронины ускоряют белковый синтез, стимулируют процессы роста и клеточной дифференцировки, ускоряют транскрипцию гена гормона роста.
В печени йодтиронины ускоряют гликолиз, синтез холестерола и синтез жёлчных кислот. В печени и жировой ткани Т3повышает чувствительность клеток к действию адреналина и косвенно стимулирует липолиз в жировой ткани и мобилизацию гликогена в печени. Т3увеличивает в мышцах потребление глюкозы, стимулирует синтез белков и увеличение мышечной массы, повышает чувствительность мышечных клеток к действию адреналина.
Йодтиронины стимулируют работу Na+,K+-ATФазы, повышают поглощение клетками кислорода (кроме мозга, РЭС и гонад).
Йодтиронины участвуют в формировании ответной реакции на охлаждение увеличением теплопродукции, повышая чувствительность симпатической нервной системы к норадреналину и стимулируя секрецию норадреналина.
Очень высокие концентрации Т3тормозят синтез белков и стимулируют катаболические процессы.
6. Инактивацияйодтиронинов осуществляется в периферических тканях в результате дейодирования Т4 до «реверсивной» Т3по 5, полного дейодирования, дезаминирования или декарбоксилирования. Йодированные продукты катаболизма йодтиронинов конъюгируют в печени с глюкуроновой или серной кислотами, секретируются с жёлчью, в кишечнике вновь всасываются, дейодируются в почках и выделяются с мочой. Для Т4Т½ =7 дней, для Т3Т½ =1-1,5 дня.
Регуляция синтеза и секреции йодтиронинов
Синтез и секреция йодтиронинов регулируется гипоталамо-гипофизарной системой.

Заболевания щитовидной железы
Гипотиреозразвивается вследствие дефицита йодтиронинов при недостаточности функции щитовидной железы (хронический аутоиммунный тиреоидит - зоб Хашимото), при заболеваниях гипофиза и гипоталамуса, при дефиците йода в пище (эндемический зоб).
Гипотериоз приводитк снижению основного обмена, скорости гликолиза, мобилизации гликогена и жиров, потребления глюкозы мышцами, уменьшения мышечной массы и снижения теплопродукции.
Проявления гипотериоза: снижение частоты сердечных сокращений, вялость, сонливость, непереносимость холода, сухость кожи.
Гипотиреоз новорождённыхприводит к развитию кретинизма (тяжёлой необратимой задержкой умственного развития). У детей старшего возраста наблюдают отставание в росте без задержки умственного развития.
Тяжёлой формой гипотиреоза является «микседема».Она сопровождается отёком кожи и подкожной клетчатки. Отёк обусловлен накоплением в межклеточном матриксе ГАГ (глюкуроновая и в меньшей степени хондроитинсерная кислоты). Избыток ГАГ
Эндемический зоб(нетоксический зоб) часто встречается у людей, живущих в районах, где содержание йода в воде и почве недостаточно. Если поступление йода в организм снижается (ниже 100 мкг/сут), то уменьшается продукция йодтиронинов, что приводит к усилению секреции ТТГ (из-за ослабления действия йодтиронинов на гипофиз по механизму отрицательной обратной связи), под влиянием которого происходит компенсаторное увеличение размеров щитовидной железы (гиперплазия), но продукция йодтиронинов при этом не увеличивается.
Гипертиреозвозникает при повышенной продукции йодтиронинов вследствие развития опухоли, тиреоидите, избыточном поступлении йода и йодсодержащих препаратов, аутоиммунных реакций.
Диффузный токсический зоб (базедова болезнь, болезнь Грейвса) — наиболее распространённое заболевание щитовидной железы. При этом заболевании отмечают увеличение размеров щитовидной железы (зоб), повышение концентрации йодтиронинов в 2—5 раз и развитие тиреотоксикоза.
Болезнь Грейвсавозникает в результате образования антител к тиреоидным антигенам. Один из них, иммуноглобулин (IgG), имитирует действие тиреотропина, взаимодействуя с рецепторами тиреотропина на мембране клеток щитовидной железы. Это приводит к диффузному разрастанию щитовидной железы и избыточной неконтролируемой продукции Т3и Т4, поскольку образование IgG не регулируется по механизму обратной связи. Уровень ТТГ при этом заболевании снижен вследствие подавления функции гипофиза высокими концентрациями йодтиронинов.
Гипертериоз приводитк увеличению основного обмена: одновременно стимулируется анаболизм (рост и дифференцировка тканей) и в большей степени катаболизм (углеводов, липидов и белков), возникает отрицательный азотистый баланс.
Проявления гипертериоза: учащение сердцебиения, мышечная слабость, снижение массы тела (несмотря на повышенный аппетит), потливость, повышение температуры тела, тремор и экзофтальм (пучеглазие).