

Трансмембранная передача информации с участием аденилатциклазной системы
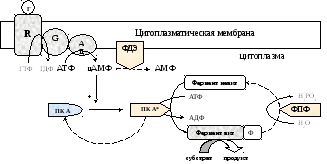
Последовательность событий, приводящих к каталитической активации ферментов
1). 1 Гормон (Г) присоединяется к Rs-рецептору с образованием гормон-рецепторного комплекса, который через несколькоGs-белков активирует несколько аденилатциклаз (комплекс гормон-Ri-рецептор черезGi-белки ингибирует аденилатциклазы);
2). 1 Аденилатциклаза превращает тысячи АТФ в тысячи цАМФ;
3). 4 цАМФ обратимо присоединяясь к 2 рецепторным субъединицам RПК А, вызывают диссоциацию протомеров ПК А с освобождением 2 активных каталитических субъединиц С и двух 2цАМФ-R;
4). 1 Субъединица С ПК А фосфорилирует сотни ферментов, что способствует их активации или ингибированию. Таким образом, сигнал 1 гормона в аденилатциклазной системе суммарно усиливается в 106-107 раз;
5). Миллионы активных ферментов превращают молекулы субстрата в продукты.
Инактивация аденилатциклазной системы осуществляется через ФДЭ и фосфотазу. ФДЭ разрушает цАМФ до АМФ, это приводит к самосборке субъединиц ПК А и ее инактивации. Фосфотаза инактивирует (активирует) фермент в реакции гидролитического дефосфорилирования.
Посредством стимуляции аденилатциклазной системы оказывают гормональное действие кортиколиберин,кальцитонин, соматолиберин, ВИП,глюкагон,вазопрессин (черезV2рецепторы), ЛГ, ФСГ,ТТГ, хорионический гонадотропин, АКТГ,паратгормон, простагландины типа Е, D и I,адренергические катехоламины(черезb1иb2рецепторы).
Трансмембранная передача информации с участием инозитолтрифосфатной системы
субстрат продукт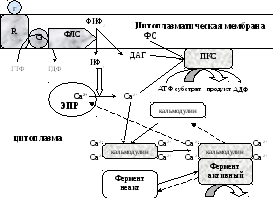
Последовательность событий, приводящих к каталитической активации ферментов
1). Гормон (Г) присоединяется к R-рецептору с образованием гормон-рецепторного комплекса, который черезG-белок активирует фосфолипазу С;
2). Фосфолипаза С расщепляет фосфатидилинозитол-4,5-дифосфат (ФИФ2) клеточной мембраны на инозитолтрифосфат (ИФ3) и диацилглицерин (ДАГ);
3). ИФ3присоединяясь к рецептору на мембране ЭПР, открывает кальциевые каналы, выпуская Са2+ из ЭПР в цитоплазму;
4). Са2+, ДАГ и фосфотидилсерин активируют ПК С, которая превращает субстрат в продукт.
5). 4 Са2+ присоединяются к кальмодулину, способствуя присоединению кальмодулина к ферменту и образованию активного комплекса 4Са2+-кальмодулин-фермент, который превращает субстрат в продукт.
Инактивация инозитолтрифосфатной системы осуществляется кальциевыми насосами, которые откачивают Са2+ из цитоплазмы.
Посредством стимуляции инозитолтрифосфатной системы оказывают гормональное действие гонадолиберин, тиролиберин, дофамин, тромбоксаны А2, эндоперекиси, лейкотриены, агниотензин II, эндотелин, паратгормон, нейропептид Y,адренергические катехоламины(через a1рецепторы), ацетилхолин, брадикинин,вазопрессин (черезV1рецепторы).
Трансмембранная передача информации с участием цитоплазматических и ядерных рецепторов
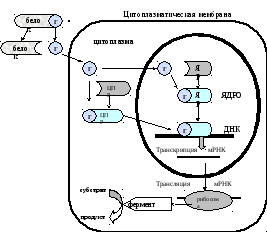
Через цитоплазматические и ядерные рецепторы действуют кортикоиды, половые и тиреоидные гормоны.
Последовательность событий, приводящих к активации транскрипции и биосинтезу ферментов:
1). Освобождаясь от белка, гормон самостоятельно проходит клеточную мембрану и в цитоплазме присоединяется к цитоплазматическому рецептору с образованием гормон-рецепторного комплекса;
2). Гормон-рецепторный комплекс мигрирует в ядро, где присоединяется к регуляторной нуклеотидной последовательности в ДНК – энхансером или сайленсером.
3). При взаимодействии с энхансером увеличивается, а с сайленсером уменьшается доступность промотора для РНК-полимеразы, соответственно меняется скорость транскрипции мРНК и трансляции новых ферментов;
4). Ферменты превращают субстраты в продукты.
Иногда, гормон самостоятельно проникает в ядро, где соединяется с ядерным рецептором, образуя гормон-рецепторный комплекс. Этот комплекс также присоединяется к ДНК, запускает или блокирует процесс транскрипции мРНК и трансляции новых ферментов.
Изменение количества ферментов и интенсивности метаболизма под действием стероидных и териоидных гормонов происходит в течение нескольких часов.