

Регуляция сокращения и расслабления мышц
Мышечное сокращение находится под сложным регуляторным влиянием со стороны нервной системы. Мышечное сокращение опосредуется Са2+.
Кальциевые насосы постоянно перекачивают Са2+ из саркоплазмы в саркоплазматический ретикулум (у скелетных мышц) или межклеточный матрикс (миокард) (при участии Са-связывающего белка - кальсеквестрина). В результате в саркоплазме покоящейся мышцы концентрация Са2+ составляет всего 10-7-10-8моль/л.
При действии, например, ацетилхолина на ацетилхолиновые рецепторы происходит возбуждение сарколеммы.
Потенциал действия сарколеммы, через Т-систему у скелетных мышц или напрямую у миокарда и гладких мышц,достигает кальциевых каналов саркоплазматического ретикулума (рианодиновые рецепторы).
Кальциевые каналы открываются, выпуская Са2+из саркоплазматического ретикулума в саркоплазму, так что его концентрация в ней возрастает до 10-5моль/л.
Далее механизм регуляции мышечного сокращения в поперечнополосатых и гладких мышцах отличается.
Актиновая регуляция
Актиновая регуляция характерна для поперечнополосатых мышц - скелетных и сердечной.
Мышечное сокращение скелетных мышц ингибирует тропомиозиновая система на 2 стадии сокращения, так как TпIпредотвращает присоединение миозиновой головки к соответствующему связывающему сайту F-актина (TпIили изменяет конформацию F-актина или перемещает тропомиозин в то положение, в котором он блокирует сайты связывания миозиновых головок на F-актине).
Поступающий в саркоплазму Са2+ присоединяется к тропонину ТnС. Комплекс ТnС•Са2+реагирует с TnIи ТnТ, влияя на их взаимодействие с тропомиозином.
Тропомиозин при этом либо отсоединяется, либо изменяет конформацию F-актина таким образом, что появляется возможность присоединения к нему миозиновой головки тяжелой цепи. Начинается сократительный цикл.
Расслабление происходит, когда 1) содержание Са2+в саркоплазме падает ниже 10-7моль/л вследствие его поглощения саркоплазматическим ретикулумом; 2) комплекс ТnС•Са2+утрачивает свой Са2+; 3) тропонин, реагируя с тропомиозином, ингибирует дальнейшее взаимодействие миозиновой головки с F-актином и 4) миозиновые головки в присутствии АТФ отделяются от F-актина, вызывая расслабление.
Так как в сердечной мышце основным источником ионов Са2+для возбуждения служит внеклеточная жидкость, при отсутствии Са2+во внеклеточной жидкости сокращения сердечной мышцы прекращаются в течение одной минуты. Скелетная мышца в таких условиях может сокращаться часами.
Исчезновение АТФ из саркоплазмы приводит к следующим последствиям: 1) Са2+-насос саркоплазматического ретикулума перестает поддерживать низкую концентрацию Са2+в саркоплазме; при этом стимулируется взаимодействие миозиновых головок с F-актином; 2) не происходит зависимого от АТФ отделения миозиновых головок от F-актина, при этом на 5 стадии мышечного сокращения наступает трупное окоченение.
Миозиновая регуляция
Миозиновая регуляция характерна для гладких мышц.
У гладких мышц нет тропониновой системы, а легкая цепь (р-цепь) миозина подавляет его АТФ-азную активность и препятствует присоединению миозина к F-актину.
В саркоплазме гладких мышц присутствует киназа легких цепей миозина, зависимая от Са2+. При повышении в саркоплазме Са2+, он присоединяется к кальмодулину. Комплекс кальмодулин-4Са2+ активирует киназу легких цепей миозина.
Активная киназа легких цепей миозина фосфорилирует легкую цепь р, которая при этом перестает ингибировать АТФ-азную активность миозина и препятствовать взаимодействию миозина с F-актином. В результате начинается сократительный цикл.
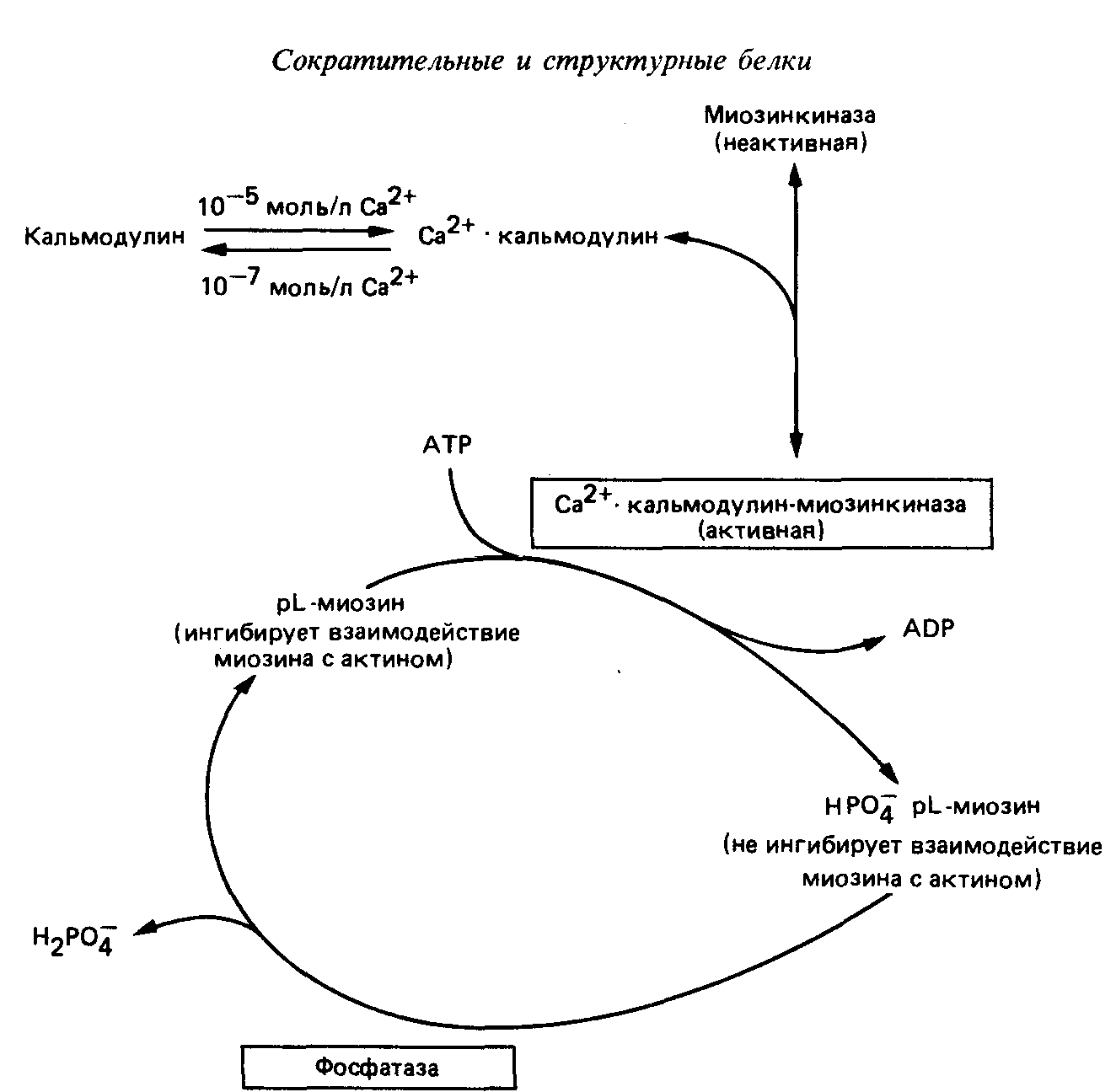
Кальциевая регуляция сокращения гладких мышц (по Р. Марри, 1993).
Расслабление гладких мышц происходит, когда 1) содержание ионов Са2+в саркоплазме падает ниже 10-7моль/л; 2) Са2+отсоединяется от кальмодулина, который в свою очередь отделяется от киназы легкой цепи миозина, вызывая ее инактивацию; 3) нового фосфорилирования легкой цепи р не происходит, и протеинфосфатаза легкой цепи, которая постоянно активна и не зависит от кальция, отщепляет от легкой цепи р ранее присоединившиеся к ней фосфаты; 4) дефосфорилированная легкая цепь р миозина ингибирует связывание миозиновых головок с F-актином и подавляет активность АТФ-азы; 5) миозиновые головки в присутствии ATФ отделяются от F-актина, а повторное их связывание произойти не может из-за присутствия в системе дефосфорилированной легкой цепи р.