

Перекисное окисление липидов
Понятие ПОЛобъединяет все реакции неферментативного окисления полиненасыщенных ЖК, свободных или входящих в состав омыляемых липидов, протекающих по радикальному механизму. Реакции ПОЛ инициируются активными формами кислорода.


В результате появления в гидрофобном слое мембран гидрофильных зон за счёт образования гидропероксидов жирных кислот в клетки могут проникать вода, ионы натрия, кальция, что приводит к набуханию клеток, органелл и их разрушению.
Регуляция ПОЛ
Процессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии (при реоксигенации), повышенном содержании активных форм О2, снижении антиоксидантной защиты, повышенном содержании ненасыщенных жирных кислот.
Активация ПОЛ происходит при воспалении и характерна для многих заболеваний: дистрофии мышц (болезнь Дюшенна), болезни Паркинсона, атеросклерозе, развитии опухолей.
Физиологическое значение реакций ПОЛ:
модифицируют физико-химические свойства биомембран: увеличивают их проницаемость.
регулируют активность мембранных ферментов, реакции окислительного фосфорилирования.
участвуют в контроле клеточного деления.
Участвует в адаптации организма.
Повышение активности ПОЛ приводит к:
разрушению, фрагментации клеточных мембран, повреждению и гибели клеток.
модификации ЛП, особенно ЛПНП. Они становятся «липкими», легче проникают в сосудистую стенку, хорошо захватываются макрофагами, что ускоряет развитие атеросклероза.
накоплению продуктов ПОЛ, многие из которых токсичны, канцерогенены и мутагенены (МДА).
ускорению процессов старения организма.
На коже с возрастом увеличивается количество пигментных пятен. Пигмент, вызывающий образование этих пятен, называется липофусцин. Он представляет собой смесь липидов и белков, связанных между собой поперечными ковалентными связями и денатурированными в результате взаимодействия с продуктами ПОЛ. Этот пигмент фагоцитируется, но не гидролизуется ферментами лизосом, и поэтому накапливается в клетках, нарушая их функции.
Анаболизм жирных кислот
Источником ЖК в организме являются синтетические процессы и пища.
ЖК, которые синтезируются в организме, называются заменимыми. Значительная их часть образуется в печени, в, меньшей степени — в жировой ткани и лактирующей молочной железе.
ЖК, которые не синтезируются в организме, но необходимы для него называются незаменимыми. Единственным источником незаменимых ЖК является пища.
У человека синтез ЖК начинается с образования пальмитиновой кислоты, из которой затем образуются другие заменимые ЖК. Кроме того, некоторые заменимые ЖК образуются из незаменимых ЖК. Субстратами для синтеза ЖК служит ацетил-КоА и НАДФН2, образующийся в основном из глюкозы. Таким образом, избыток углеводов, поступающих в организм, трансформируется в ЖК, а затем в ТГ.
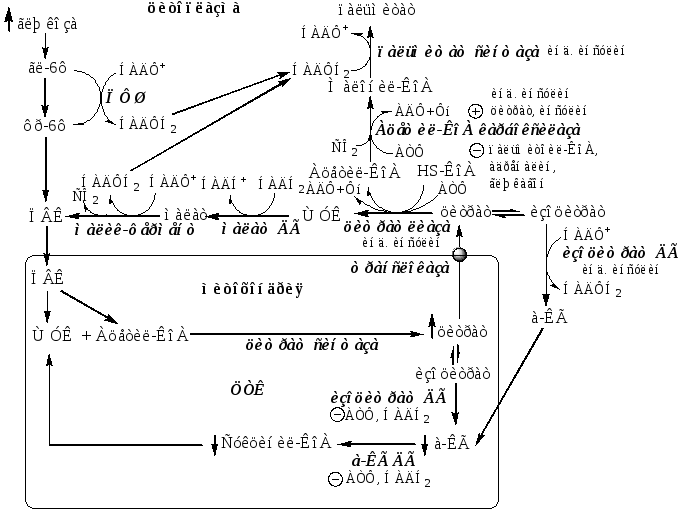
Образование субстратов, необходимых для синтеза ЖК
Образование и транспорт Ацетил-КоА. В реакциях гликолиза из глюкозы образуется ПВК, который поступает в матрикс митохондрий и превращается в Ацетил-КоА с участием ПВК ДГ. Так как внутренняя мембрана митохондрий непроницаема для Ацетил-КоА, поэтому он при участии цитратсинтазы конденсируется с ЩУК с образованием цитрата:
Ацетил-КоА + Оксалоацетат → Цитрат + HS-КоА.
Затем транслоказа переносит цитрат в цитоплазму. Перенос цитрата в цитоплазму происходит только при увеличении количества цитрата в митохондриях, когда изоцитратдегидрогеназа и α-кетоглутаратдегидрогеназа ингибированы высокими концентрациями НАДН2и АТФ (при избытке углеводов и низком энергопотреблении).
В цитоплазме цитрат расщепляется под действием фермента цитрат-лиазы:
Цитрат + HSKoA + АТФ → Ацетил-КоА + АДФ+ Pн + ЩУК
Образование НАДФН2.
ЩУК в цитоплазме превращается в малат под действием малат ДГ, малат под действием малик-фермента превращается в ПВК, при этом образуется НАДФН2. ПВК транспортируется обратно в матрикс митохондрий;
НАДФН2 образуется в цитоплазме из глюкозы в окислительных реакциях ПФШ;
Цитрат изомеризуется в изоцитрат, который дегидрируется цитозольной НАДФ-зависимой ДГ до α-КГ с образованием НАДФН2. α-КГ переноситься в матрикс митохондрий.
 Синтез
пальмитиновой кислоты
Синтез
пальмитиновой кислоты
Образование малонил-КоА
Первая реакция синтеза ЖК — превращение ацетил-КоА в малонил-КоА. Это регуляторная реакция в синтезе ЖК катализируется ацетил-КоА-карбоксилазой.
Ацетил-КоА-карбоксилаза состоит из нескольких субъединиц, содержащих биотин.
Реакция протекает в 2 стадии:
СО2 + биотин + АТФ → биотин-СООН + АДФ + Фн
ацетил-КоА + биотин-СООН → малонил-КоА + биотин
Ацетил-КоА-карбоксилаза регулируется несколькими способами:
Ассоциация/диссоциация комплексов субъединиц фермента. В неактивной форме ацетил-КоА-карбоксилаза представляет собой комплексы, состоящих из 4 субъединиц. Цитрат стимулирует объединение комплексов, в результате чего активность фермента увеличивается. Пальмитоил-КоА вызывает диссоциацию комплексов и снижение активности фермента;
Фосфорилирование/дефосфорилирование ацетил-КоА-карбоксилазы. Глюкагон или адреналин через аденилатциклазную систему стимулируют фосфорилирование субъединиц ацетил-КоА карбоксилазы, что приводит к ее инактивации. Инсулин активирует фосфопротеинфосфатазу, ацетил-КоА карбоксилаза дефосфорилируется. Затем под действием цитрата происходит полимеризация протомеров фермента, и он становится активным;
Длительное потребление богатой углеводами и бедной липидами пищи приводит к увеличению секреции инсулина, который индукцирует синтез ацетил-КоА-карбоксилазы, пальмитатсинтазы, цитратлиазы, изоцитратдегидрогеназы и ускоряет синтез ЖК и ТГ. Голодание или богатая жирами пища приводит к снижению синтеза ферментов и, соответственно, ЖК и ТГ.
Образование пальмитиновой кислоты
После образования малонил-КоА синтез пальмитиновой кислоты продолжается на мультиферментном комплексе — синтазе жирных кислот (пальмитоилсинтетазе).
Пальмитоилсинтаза - это димер, состоящий из двух идентичных полипептидных цепей. Каждая цепь имеет 7 активных центров и ацилпереносящий белок (АПБ). В каждой цепи есть 2 SH-гpyппы: одна SH-гpyппa принадлежит цистеину, другая — остатку фосфопантетеиновой кислоты. SH-группа цистеина одного мономера расположена рядом с SH-группой 4-фосфопантетеината другого протомера. Таким образом, протомеры фермента расположены «голова к хвосту». Хотя каждый мономер содержит все каталитические центры, функционально активен комплекс из 2 протомеров. Поэтому реально синтезируются одновременно 2 ЖК.
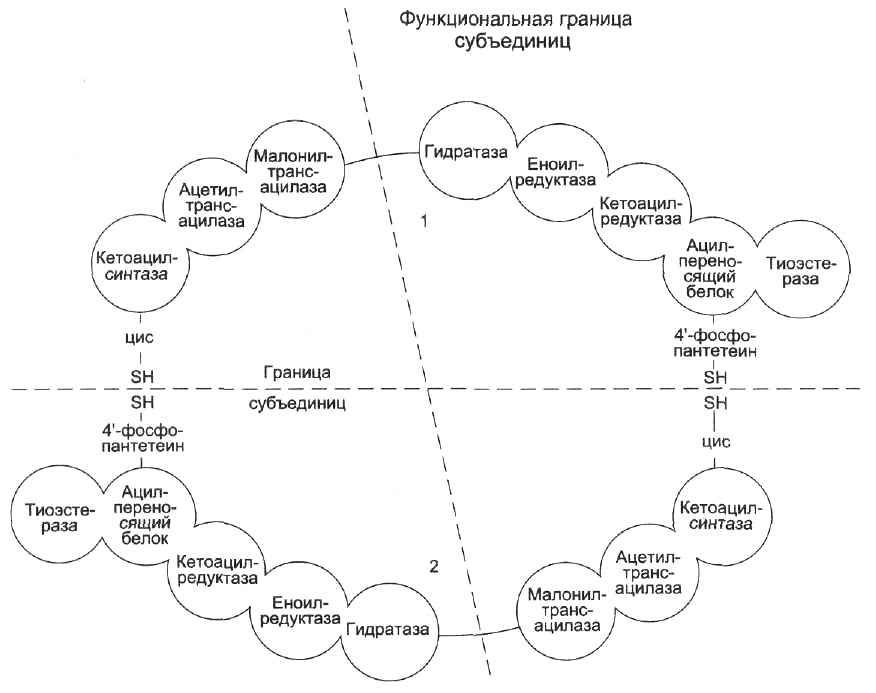
Этот комплекс последовательно удлиняет радикал ЖК на 2 атома С, донором которых служит малонил-КоА.
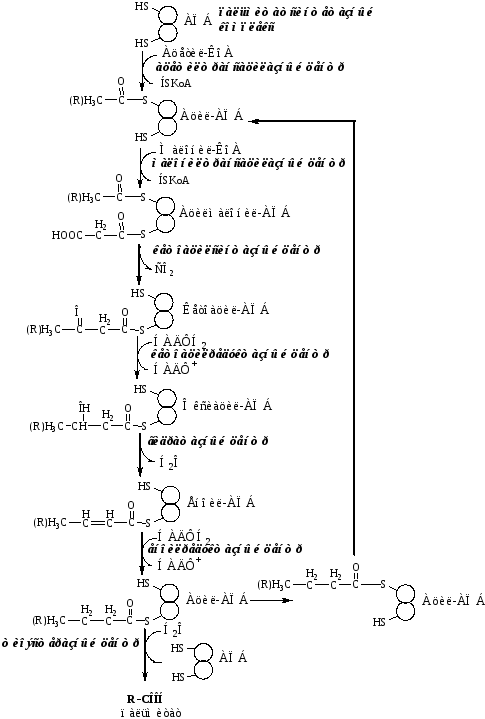
Реакции синтеза пальмитиновой кислоты
Перенос ацетила с КоА на SH-группу цистеина ацетилтрансацилазным центром;
Перенос малонила с КоА на SH-группу АПБ малонилтрансацилазным центром;
Кетоацилсинтазным центром ацетильная группа конденсируется с малонильной с образованием кетоацила и выделением СО2.
Кетоацил восстанавливается кетоацил-редуктазой до оксиацила;
Оксиацил дегидратируется гидратазой в еноил;
Еноил восстанавливается еноилредуктазой до ацила.
В результате первого цикла реакций образуется ацил с 4 атомами С (бутирил). Далее бутирил переносится из позиции 2 в позицию 1 (где находился ацетил в начале первого цикла реакций). Затем бутирил подвергается тем же превращениям и удлиняется на 2 атома С (от малонил-КоА).
Аналогичные циклы реакций повторяются до тех пор, пока не образуется радикал пальмитиновой кислоты, который под действием тиоэстеразного центра гидролитически отделяется от ферментного комплекса, превращаясь в свободную пальмитиновую кислоту.
Суммарное уравнение синтеза пальмитиновой кислоты из ацетил-КоА и малонил-КоА имеет следующий вид:
CH3-CO-SKoA + 7 HOOC-CH2-CO-SKoA + 14 НАДФН2 → C15H31COOH + 7 СО2 + 6
Н2О + 8 HSKoA + 14 НАДФ+
Синтез ЖК из пальмитиновой и других ЖК
Удлинение ЖК в элонгазных реакциях
Удлинение ЖК называется элонгацией. ЖК могут синтезироваться в результате удлинение в ЭПР пальмитиновой кислоты и других более длинных ЖК. Для каждой длины ЖК существуют свои элонгазы. Последовательность реакций аналогична синтезу пальмитиновой кислоты, однако в данном случае синтез идет не на АПБ, а на КоА. Основной продукт элонгации в печени — стеариновая кислота. В нервных тканях образуются ЖК с длинной цепью (С=20-24), необходимые для синтеза сфинголипидов.
Синтез ненасыщенных ЖК в десатуразных реакциях
Включение двойных связей в радикалы ЖК называется десатурацией. Десатурация ЖК происходит в ЭПР в монооксигеназных реакциях, катализируемых десатуразами.
Стеароил-КоА-десатураза– интегральный фермент, содержит негеминовое железо. Катализирует образование 1 двойной связи между 9 и 10 атомами углерода в ЖК. Стеароил-КоА-десатураза переносит электроны с цитохромаb5на 1 атом кислород, при участии протонов этот кислород образует воду. Второй атом кислорода включается стеариновую кислоту с образованием её оксиацила, который дегидрируется до олеиновой кислоты.
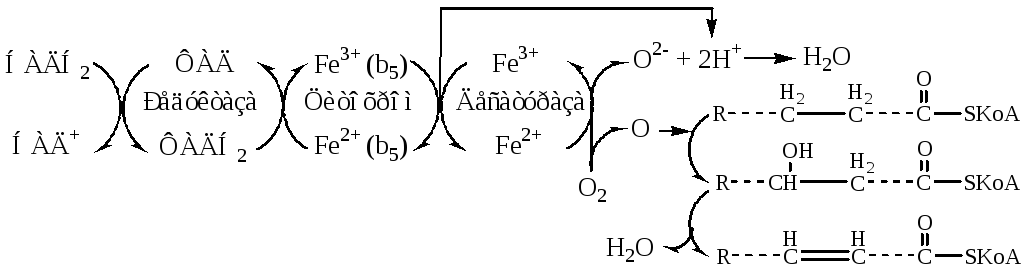
Десатуразы ЖК, имеющиеся в организме человека, не могут образовывать двойные связи в ЖК дистальнее девятого атома углерода, поэтому ЖК семейства ω-3 и ω-6 не синтезируются в организме, являются незаменимыми и обязательно должны поступать с пищей, так как выполняют важные регуляторные функции. Основные ЖК, образующиеся в организме человека в результате десатурации — пальмитоолеиновая и олеиновая.
Синтез α-гидрокси ЖК
В нервной ткани происходит синтез и других ЖК — α-гидроксикислот. Оксидазы со смешанными функциями гидроксилируют С22и С24кислоты с образованием цереброновой кислоты обнаруживаемой только в липидах мозга.