

24. Возрастные особенности в количестве лейкоцитов. Двойной перекрест в соотношении нейтрофилов и лимфоцитов у детей.
Количество лейкоцитов у новорожденных повышено и равно 10- 30 *109 /л. Число нейтрофилов составляет -60,5 %, эозинофилов – 2%, базофилов -02 %, моноцитов -1,8 %, лимфоцитов – 24 %. В течении первых 2 недель количество лейкоцитов сокращается до 9 – 15 *109 /л, к 4 годам уменьшается до 7-13*109 /л, а к 14 годам достигает уровня, характерного для взрослого. Соотношение нейтрофилов и лимфоцитов меняется, что обуславливает возникновение физиологических перекрестов.
Первый перекрест. У новорожденного соотношение содержания этих клеток такое же, как и у взрослого. В последующем сод. Нф падает, а Лмф возрастает, так что на 3-4 сутки их количество уравнивается. В дальнейшем количество Нф продолжает снижаться и к 1-2 годам достигает 25 %. В этом же возрасте количество Лмф- 65 %.
Второй перекрест. В течении следующих лет число Нф постепенно повышается, а Лмф –снижается, так что у детей в 4 года эти показатели снова уравниваются и составляют по 35 % от общего количества лейкоцитов. Количество Нф продолжает увеличиваться, а количество Лмф – уменьшается, и к 14 годам эти показатели соответствуют таковым у взрослого (4-9 *109 /л).
25. Генез, структура, общие и спец. Свойства и функции нейтофилов
В костном мозге можно наблюдать шесть последовательных морфологических стадий созревания нейтрофилов: миелобласт, промйелоцит, миелоцит, метамиелоцит, палочкоядерная и сегментоядерная клетка:
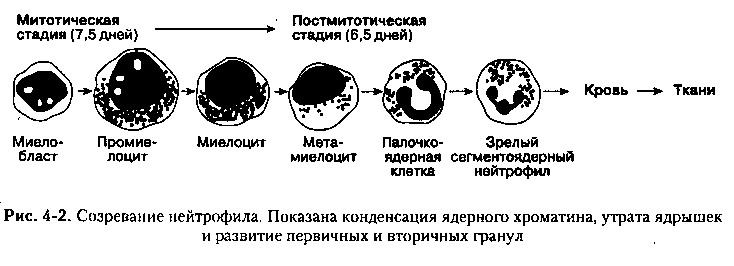
Кроме того, там же имеются более ранние, морфологически не идентифицируемые, коммитированные предшественники нейтрофилов: КОЕ-ГМ и КОЕ-Г.
Созревание нейтрофилов сопровождается прогрессирующим снижением размера ядра за счет конденсации хроматина и потери ядрышек. По мере созревания нейтрофила ядро зазубривается и наконец приобретает характерную сегментацию. Одновременно происходят изменения и в цитоплазме нейтрофила, где накапливаются гранулы, содержащие биологические соединения, которые впоследствии будут играть столь важную роль в защите организма. Первичные (азурофильные) гранулы — включения синего цвета размером приблизительно 0,3 мкм, содержащие эластазу и миелопероксидазу. Впервые они появляются на промиелоцитарной стадии; при созревании их количество и интенсивность окрашивания снижаются. Вторичные (специфические) гранулы, которые содержат лизоцим и другие протеазы, появляются на стадии миелоцита. Окраска этих вторичных гранул обусловливает характерный нейтрофильный вид цитоплазмы.
Кинетика нейтрофилов. По способности к делению миелобласты, промиелоциты и миелоциты относятся к митотической группе, т.е. обладают способностью к делению, интенсивность которого падает от миелобласта к миелоциту. Последующие этапы созревания нейтрофилов не связаны с делением. В костном мозге пролиферирующие клетки среди нейтрофилов составляют около 1/3, и столько же приходится на долю гранулоцитарных митозов среди всех пролиферирующих клеток костного мозга. В течение суток вырабатывается до 4,0x109 нейтрофилов на килограмм массы тела.
Структура. Цитоплазма нейтрофилов. На стадии метамиелоцита и последующих стадиях созревания редуцируются структуры, обеспечивающие синтез цитоплазматических белков, совершенствуется структура лизосом, обеспечивающих функцию нейтрофилов, усиливается способность к амебовидной подвижности, деформации, обеспечивающих подвижность и инвазивность гранулоцитов.
Мембрана нейтрофилов. На предшественниках гранулоцитарного ростка определяются CD34+CD33+, а также рецепторы для G M - C S F , G - C S F, IL-1 , IL-3, IL-6, IL-11 , IL-12. На мембране присутствуют также различные молекулы, являющиеся рецепторами для хемотаксических сигналов, к которым относятся CCF , N-формил-пептид.
Свойства и функции. Функция нейтрофилов заключается в защите организма от инфекции. Этот процесс включает хемотаксис, фагоцитоз и уничтожение микроогранизмов. Хемотаксис предполагает способность к обнаружению и целенаправленному движению по направлению к микроорганизмам и очагам воспаления. Нейтрофилы имеют специфические рецепторы для С5а-компонента системы комплемента (вырабатываемого в классическом или альтернативном путях активации комплемента) и протеаз, выделяемых при повреждении тканей или при непосредственном бактериальном воздействии. Кроме того, у нейтрофилов есть рецепторы для N-формилъных пептидов, выделяемых бактериями и пораженными митохондриями. Они реагируют и на такие продукты воспаления, лейкотриен LТВ-4 и фибринопептиды.
Нейтрофилы распознают инородные организмы при помощи рецепторов к опсонинам. Фиксация сывороточного IgG и комплемента на бактериях делает их распознаваемыми для гранулоцитов. Нейтрофил имеет рецепторы для Fc-фрагмента молекулы иммуноглобулина и продуктов каскада комплемента. Эти рецепторы инициируют процессы захвата, поглощения и адгезии инородных объектов.
Нейтрофилы поглощают опсонизированные микроорганизмы с помощью цитоплазматических пузырьков, называемых фагосомами. Эти пузырьки продвигаются от складчатых псевдоподий и сливаются с первичными и вторичными гранулами за счет энергетически зависимого процесса, во время которого в фагоцитах происходит взрывная активация гликолиза и гликогенолиза. При дегрануляции клетки содержимое гранул выбрасывается в фагосому и выделяются ферменты деградации: лизоцим, кислая и щелочная фосфатазы, эластазаилактоферрин.
Наконец, нейтрофилы разрушают бактерии, метаболизируя кислород с образованием продуктов, токсичных для поглощенных микроорганизмов. Оксидазный комплекс, генерирующий эти продукты, состоит из флавин- и гемсодержащегося цитохрома Ь558-.
В этих реакциях используется восстанавливающий агент НАДФН, а стимуляторами их являются глюкозо-6-фосфатдегидрогеназа и другие ферменты гексозомонофосфатного шунта. В результате клетка генерирует супероксид (О2) и перекись водорода (Н202), которые выделяются в фагосому для уничтожения бактерий. Лактоферрин участвует в образовании свободных гидроксильных радикалов■, а миелопероксидаза, используя галоиды в качестве кофакторов,— в продукции гипохлорной кислоты (НОС1) и токсичных хлораминов.