

19. Компоненты этц и нммк фотосистем 1 и 2, их редокс-потенциалы
Пластоцианин — маленький, подвижный белок с молекулярной массой около 10,5 кДа. К его центральному атому Сu присоединяются остатки цистеина и метионина, а сбоку его стабилизируют два остатка гистидина. При обратимой смене валентности Cu2+ → Cu+1 пластоцианин либо поглощает один электрон, либо отдаёт его. Пластоцианин является аналогом цитохрома c, который выполняет схожую функцию в дыхательной цепи митохондрий. Он принимает электрон от цитохром b6f-комплекса, окисляя цитохром f и перенося его непосредственно на реакционный центр П700 фотосистемы I. На внешней стороне белка имеется группа аминокислот, несущих отрицательный заряд.
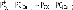
П700 - представляет собой димерхлорофилла а и хлорофилла а‘, у которого кетоэфирная группа в V кольце находится в цис-положении относительно плоскости молекулы, с максимумом поглощения 700 нм. П700 получает энергию от антенных комплексов и использует её, чтобы поднять электроны на более высокий уровень. Далее электрон в ходе окислительно-восстановительной реакции переходит на цепь переносчиков. В окисленном состоянии окислительно-восстановительный редокс-потенциал П700 составляет +0,52 В, а в фотовозбуждённом состоянии он становится −1,2 В, то есть формируется мощный восстановитель, обеспечивающий восстановление НАДФ+.
Хлорофилл А0— первый акцептор электронов в фотосистеме I. Именно здесь происходит первичное фотохимическое разделение зарядов между фотовозбуждённым П700* и A0. Его максимум поглощения составляет 695 нм (Хл а695), что объясняется его взаимодействием с окружающими аминокислотными остатками. Его редокс-потенциал в восстановленном состоянии −1,1 В[1]. Фотовозбуждённый П700* отдаёт один электрон хлорофиллу A0, в результате чего происходит разделение зарядов, и образуется первичная радикальная пара:

Филлохинон A1 также известный как витамин K1. Он, как и хлорофилл, обладает фитольным хвостом, и приблизительно соответствует пластохинону QA фотосистемы II. Поглощая электрон, он образует семихинон-радикал, который восстанавливает Fx, передаёт его на Fb и далее на Fa.

Железосерные кластеры ФСI имеют форму куба с четырьмя атомами железа и четырьмя атомами серы, составляющими его восемь вершин. Все три кластера связаны с белками ФСI через остатки цистеина. Fx окисляет восстановленный А1. Дальнейший транспорт осуществляют железосерные кластеры Fa, и Fb, характеризующиеся низкими окислительно-восстановительными потенциалами. Fx, Fa, и Fb образуют треугольник, и Fa располагается ближе к Fx чем Fb. Транспорт электронов начинается с Fx через Fa к Fb, или же через Fa к Fb.
Ферредоксин — это водорастворимый белок с молекулярной массой 11 кДа и содержащий Fe2S2 центр. Одноэлектронная окислительно-восстановительная система, то есть он переносит только один электрон, полученный от железосерных кластеров. Он восстанавливается ФСI на стромальной стороне мембраны и в восстановленном состоянии является сильным восстановителем, благодаря чему может быть переносчиком электронов для различных реакций, протекающих в хлоропласте. ТакПри нециклическом транспорте электронов ферредоксин взаимодействует с ферредоксин-НАДФ+ редуктазой, которая восстанавливает НАДФ+ до НАДФН в строме хлоропласта.
Tyrz — остаток тирозина белка D1 (Tyr-161). Это промежуточный переносчик электронов, который переносит электроны между марганцевым кластером и П680. Перенос электронов происходит с образованием нейтрального радикала.

П680— это пара хлорофиллов a, с максимумом поглощения на длине волны в 680 нм. Поглощая энергию света, она отдаёт один электрон на феофитин, а сама окисляется и становится сильным окислителем П680+ с окислительно-восстановительным потенциалом +1,12 В[16], что позволяет ей индуцировать процесс окисления воды, окислительно-восстановительный потенциал которой +0,8 В. В то же время редокс-потенциал фотовозбуждённого П680 находится в отрицательной области (менее —0,6 В). Из-за возникновения локального электрического поля происходит изменение конформации специальной пары, что облегчает дальнейший перенос электрона на феофитин, а положительный заряд локализуется на одном из хлорофиллов. П680 поглощает квант света и переходит в фотовозбуждённое состояние, в результате чего один из его электронов переходит с основного подуровня S0 на первый синглетный подуровень S1:

Феофитин — первый акцептор электронов в фотосистеме II. Именно здесь, между феофитином (Eо‘ = —0,53 В) и фотовозбуждённым пигментом П680, происходит первичное фотохимическое разделение зарядов. Перенос электрона осуществляется в течение нескольких пикосекунд. Фотовозбуждённый П680* отдаёт один электрон феофитину, в результате чего происходит разделение зарядов, и образуется первичная радикальная пара:
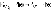
Пластохиноны QA и QBОба пластохинона играют роль вторичных акцепторов электрона, принимая его от феофитина. Перенос электрона между феофитином и пластохиноном происходит в первые 200 пикосекунд.
Цитохром b559
Цитохром b559 — гетеродимерный белок, состоящий из одной альфа (PsbE) и одной бета (PsbF) субъединицы, между которыми расположен гем. Этот белок является одним из основных компонентов ядра фотосиcтемы II. Обнаружены две формы цитохрома b559: высокопотенциальная (b559H Eо‘ = +0,37 В) и низкопотенциальная (b559L Eо‘ = +0,08 В). Высокопотенциальная форма протонирована, низкопотенциальная — депротонирована.