

1 СЕМЕСТР. Экономика. Микроэкономика. Поведение, институты и эволюция Самуэль Боулз / Микроэкономика. Поведение, институты и эволюция_Самуэль Боулз_2010 -576с
.pdfГлава 2. Спонтанный порядок: самоорганизация экономической жизни 59
Первая из них, случайность, играет центральную роль в эволюционной ди намике, даже когда стохастические события незначительны или редки. Случай
ные события могут принимать форму наследуемой инновации (как в случае мутаций). Случайность может также стать результатом поведенческих инноваций,
не являющихся (как и мутации) наилучшими ответами. В противоположность мутациям, поведенческие инновации не передаются генетически. Наоборот, они могут быть переданы и скопированы следующим поколением посредством культурного переноса, т. е. через обучение адаптивных агентов. Еще один путь,
через который случай влияет на эволюционную динамику — так называемое искажением при подборе пары. Когда небольшое число людей в неоднородной
популяции случайным образом находит себе пару для взаимодействия, реальное распределение типов людей, с которыми человек попадает в пары в течение не
которого периода, может существенно разниться с ожидаемым. Разница между реальным и ожидаемым распределением и отражает искажение при подборе
пары, способное иметь существенные последствия.
Никто не сомневается в том, что случайные события играют свою роль: эк зогенные шоки вследствие новых вкусов или технологий меняют равновесные цены и количество в стандартной статической модели рыночного равновесия. Чем же тогда отличаются эволюционные модели? Вопервых, очевидно, что ис точником различий становятся мутации, поведенческие инновации и искаже ния при подборе пары, потому что эти источники случайных шоков являются эндогенными в эволюционных моделях. Вовторых, в свете существования обоб щенной возрастающей отдачи незначительные маловероятные события зача стую оказывают существенное и продолжительное воздействие благодаря поло жительной обратной связи, а не нейтрализуются отрицательной.
Можно было бы подумать, что идея случайных событий способна привне сти в эволюционные модели искажения, которые сказываются только на ско рости изменений или на вопросе второго уровня значимости: наблюдаем ли мы в реальном мире именно равновесные состояния или лишь состояния, близкие к равновесным. Но это далеко не так: введение в анализ случайных событий зача стую влияет на направление (а не только на скорость) эволюционных измене ний, хотя, что удивительно, это не только не усложняет, а зачастую позволяет точнее предсказать вероятный исход (примеры я приведу ниже и в гл. 5).
Ни случайные события, ни преднамеренные инновации не являются доста точными для понимания эволюции человеческого поведения и институтов. Лишь
сочетание этих источников инноваций в совокупности со второй характеристи кой эволюционного подхода — дифференциальной репликацией (иногда назы
ваемой отбором) — и задает направление эволюционного процесса. Ключевая идея подхода состоит в том, что наблюдаемые институциональные и поведен ческие характеристики индивидов и обществ были в свое время скопированы и распространены, т. е. реплицированы, в то время как конкурирующие правила, веры и предпочтения угасли (или были реплицированы лишь в маргинальных сообществах).
С помощью представленных моделей будет показано, что дифференциальная репликация принимает много форм, объединяемых в группы по генетическим

60 Часть I. Координация и конфликт: базовые социальные взаимодействия
или культурным признакам. На уровне популяции распределение типов поведе ния, на которые влияют гены, может меняться изза быстрого распространения некоторых генотипов в ущерб другим. Распределение их меняется со временем изза случайных событий (дрейфа) и естественного отбора. В моделях, описы вающих этот процесс, платежи измеряют репродуктивный успех соответству ющих фенотипов, т. е. их приспособленность. Упростить задачу (зачастую даже без искаженных результатов) позволяет исключение подробностей генетическо го наследования и взаимодействия генотипа и фенотипа, а также рассмотрение поведения так, как если бы оно было фенотипическим выражением единствен ного гена, и изучение детерминант репродуктивного успеха этого гена (это мы проделаем позднее в модели «Ястреб — Голубь»). Переход от генов к поведению по большей части не исследован и конечно же включает незначительное количе ство (если включает вообще) простых зависимостей «ген — поведение», предпо
лагаемых данным методом.
Культурные признаки относятся к такому поведению, которое было усвоено,
а не передалось генетически от родителей. Обучение родителями иногда назы вают вертикальным культурным переносом, а учителями или другими людьми из поколения родителей — косвенным переносом; перенимание поведения от сверстников называется горизонтальным переносом. Аналог дифференциро
ванного приспособления в моделях культурной эволюции — скорость, с которой индивиды меняют свое поведение на новое. Данный дифференцированный про цесс копирования, как и генетическое наследование, изучен мало, но включает в себя тенденции приспособления поведения вследствие действия одной или не
скольких из следующих причин: изза того, что такое поведение принято в дан ной местности (конформизм, приспособляемость); изза того, что в прошлом
такое поведение приводило к получению бо2льших выигрышей по сравнению с другими линиями поведения (укрепляющее обучение), или потому, что это по
ведение максимизирует ожидаемые выигрыши при данных ожиданиях индиви да относительно поведения остальных членов популяции (корректировка наилучшей реакции или наилучшего ответа). Поскольку это просто, достоверно и
универсально, я смоделирую культурный перенос с корректировкой наилучшего ответа, иногда скомбинированный с конформистским обучением.
На процесс генетической и культурной эволюции большое влияние оказыва ет социальная структура: формирование связей внутри групп, схемы расселения, миграция и пр. Поскольку эти и многие другие аспекты социальной структуры основаны на усвоенном поведении, распространение переданных с культурой признаков в популяции может повлиять и на генетическую эволюцию. Это, а
также противоположный процесс — влияние распространения генов на куль турную эволюцию — называются генно-культурными эволюционными процес-
сами (пример модели для них я приведу в гл. 13). Хотя культурные и генетиче ские изменения определяют друг друга, существует, однако, разница в скоростях их распространения. Изменения в распределении генов выявляются лишь спу стя поколения и в ответ на достаточно редкие случайные события, в то время как восприятие культуры может принимать формы поведенческих эпидемий, как это произошло всего лишь за одну декаду 1960 г., когда во многих европейских языках резко возросло использование в обиходе обычных местоимений.

Глава 2. Спонтанный порядок: самоорганизация экономической жизни 61
Каким бы ни был процесс дифференциальной репликации — культурным или генетическим — его обычно моделируют, используя репликационное уравнение, описывающие репликационную динамику, приведенную выше. Репликаци
онная динамика предлагает альтернативу сравнительной статике и другим под ходам, в которых время явно не моделируется. Она позволяет полностью учесть внеравновесные популяционные процессы, основываясь как на эмпирически до стоверных предположениях об индивидуальных мыслительных способностях и поведении, так и на отражении подробностей социальных взаимодействий (кто с кем встретился, для чего, каковы итоговые платежи, какова информация и т.п.).
Таким образом, третьей характеристикой эволюционных подходов становится
явный учет внеравновесной динамики.
Явный анализ динамики дает два преимущества. Вопервых, это эволюционно нерелевантное равновесие. Явно заданная динамика проливает свет на
взаимосвязь между приведенными в предыдущей части концепциями поиска решений — равновесием по Нэшу и доминированием — и более полным и ус тойчивым понятием эволюционной стабильности. Мы увидим (здесь, в гл. 6 и особенно в гл. 12), что в правдоподобных моделях дифференциальной реплика ции некоторые равновесия по Нэшу могут оказаться фактически нерелевант ными тому, что происходит в реальных обществах, если принимать во внимание эволюционные процессы.
Второе преимущество моделирования динамических процессов в явном виде заключается в том, что в реальности существуют крайне важные неравновесные состояния реального функционирования экономики. Поскольку такое заявле ние бросает вызов базовым принципам привычного хода рассуждений в эконо мике, позвольте мне проиллюстрировать его при помощи эмпирического при мера. На многих рынках в течение длительного времени сосуществуют те, кого мы назвали бы победителями, и те, кого можно назвать проигравшими, что про тиворечит нашим ожиданиям относительно экономического равновесия. Среди фирм, производящих одни и те же товары и продающих их одним и тем же по требителям, например, на высококонкурентных рынках металлообрабатываю щей отрасли США в начале 1990 гг., наиболее успешные фирмы (если измерять успех производительностью труда) были более чем в три раза более производи тельными, чем наименее успешные, а четверть лучших фирм из них — более чем в два раза производительнее, чем четверть худших (Luria, 1996). Данные конца 1990 гг. по индонезийской электронной промышленности, являющейся частью высококонкурентного мирового рынка, показывают, что фирмы, входящие в
верхний квантиль, были в восемь раз более производительны, чем фирмы, вхо дящие в последний квантиль (Hallward-Driemeier, Iorossi & Sokoloff, 2001). Ко
нечно, индонезийский случай — крайность, а некоторая часть этих различий — лишь статистическая погрешность, и фирмы с высокой производительностью будут процветать, а низкопроизводительные фирмы — постепенно уходить из отрасли. Но процесс отбора, по всей видимости, достаточно слаб даже в таких конкурентных отраслях, что заставляет усомниться в пригодности предположе ния о том, что фирмы работают на границе производственных возможностей. Немедленное достижение равновесных состояний еще менее вероятно в усло

62 Часть I. Координация и конфликт: базовые социальные взаимодействия
виях, когда вход и выход в отрасль сильно ограничены и в которых агенты по сути не стремятся только зарабатывать деньги, а просто живут своей жизнью.
Игнорирование неравновесных состояний на том основании, что они вы мышлены, обычно становится плохим проводником в практических вопросах. Продолжим вышеприведенный пример: заметный вклад в окончание золотого века быстрого роста производительности в американской экономике после Вто
рой мировой войны внесло снижение степени выхода низкопроизводительных фирм с рынка (Bowles, Gordon & Weisskopf, 1983). Стремительный рост произво
дительности в шведской экономике в третьей четверти прошлого века был вы зван, в частности, перемещением рабочей силы и других ресурсов из низкопроиз водительных фирм к высокопроизводительным, чему способствовала политика
уравнивания зарплат и последовавшее за этим исчезновение фирмнеудачниц (Hibbs, 2000).
Хотя анализ внеравновесной динамики дает много нового для подобных политических вопросов, он более требователен по сравнению с привычным подходом сравнительной статики. Однако долгосрочное среднее изменение интересующих нас переменных всегда можно исследовать аналитически или сымитировать, что зачастую приводит к достаточно сильным выводам. Примеры приведем в гл. 11—13.
Четвертая характерная идея эволюционного моделирования состоит в том, что популяция структурирована иерархически и дифференциальная репликация может происходить на более чем одном уровне. Индивиды взаимодей
ствуют с индивидами, при этом они также входят в состав групп (например, семей, фирм) и других сообществ более высокого порядка (наций, этнических групп), и эти многосоставные группы тоже взаимодействуют. Получается, что индивиды — это пересечения взаимодействующих ячеек. Процесс дифферен циальной репликации обычно происходит одновременно на многих уровнях: в самих индивидах, между ними, между группами и т. д. Например, внутри фирм различные типы поведения (усердная работа или работа спустя рукава) копиру ются или отрицаются индивидами, что же касается фирм, то организационная структура наиболее прибыльных компаний копируется, наименее прибыльные же компании разоряются.
|
|
Таблица 2.1 |
|
Некоторые процессы, лежащие в основе эволюции поведения |
|||
|
|
|
|
Репликатор |
Уровень отбора |
||
|
|
||
индивидуальный |
группа индивидов |
||
|
|||
|
|
|
|
Изученное поведение |
Социальное обучение |
Подражание обычаям других |
|
|
(конформизм, лучший ответ, |
групп, культурная ассимиля |
|
|
укрепляющее обучение) |
ция неуспешных групп |
|
|
|
|
|
Гены |
Различный репродуктивный |
Биологическое угасание |
|
|
успех, дрейф |
неуспешных групп, |
|
|
|
сокращение пригодности |
|
|
|
подчиненных популяций |
|

Глава 2. Спонтанный порядок: самоорганизация экономической жизни 63
Таким образом, то, что реплицируется (или не реплицируется), может быть таким признаком индивида, как его предпочтения или веры; однако институты или другие групповые характеристики фирм, этнических групп или государств тоже могут служить предметом дифференциальной репликации. Адекватная этому теория должна освещать, каким образом в популяции формируются структуры групп, каким образом устанавливаются границы между итоговыми сообществами более высокого уровня и как они прекращают свое существова
ние. Одновременная работа разноуровневых дифференциальных репликаций, называемая многоуровневым отбором (или групповым отбором), приводит к
запуску совместного эволюционного процесса, управляющего траекториями изменения как индивидуальных, так и групповых характеристик (пример — совместная эволюция индивидуальных предпочтений и групповых структур — приводится в гл. 13).
В табл. 2.1 объединены варианты вышеназванных процессов, выделены репликаторы (копируемые признаки) и уровни отбора (на которых и идет предпо
лагаемое соревнование за успех репликации). Репликатор — то, что копируется: гены и шутки, индивидуальные предпочтения и веры, групповые соглашения и другие институты.
Объяснение поведения и институтов с точки зрения существования диф ференциальной репликации может показаться тавтологией. Действительно, дифференциальная репликация — это система счисления, бесполезная для про верки логики сложного аргумента. Но это также и аналитический каркас, поз воляющий совершить умозаключения, которые вряд ли можно получить иным путем. Конечно, для обоснования данного утверждения следует рассмотреть сам процесс репликации, чем бы он ни был: зависимым от прибыльности про цессе выживания или гибели фирм с различной организационной структурой; дифференцированной биологической приспособленностью или культурным соперничеством индивидов с различными поведенческими характеристиками; распространением или исчезновением социальных институтов в межгрупповых конфликтах или в ходе других процессов отбора.
Следующий пример прольет свет на некоторые отличительные особенности эволюционного подхода.
Сегрегация по месту жительства как эволюционный процесс
Как может ученый, изучающий эволюционные общественные науки, объяснить, почему люди предпочитают жить в смешанных районах, но в действительности
таких районов очень мало? Перед вами пример, иллюстрирующий некоторые характерные исходы эволюционного моделирования: множественность равновесий и историческую обусловленность этих исходов, систему локальной однородности и глобальной неоднородности и долгосрочного сохранения неэффективных по Парето исходов. Рассмотрим район (один из многих), в котором все
жилища обладают одинаковой привлекательностью для всех членов популяции.

64 Часть I. Координация и конфликт: базовые социальные взаимодействия
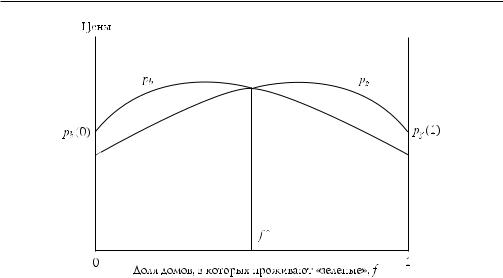
Предпочтения индивидов относительно проживания в районе зависят только от расового состава соседей. В этом районе и вокруг «зеленые» предпочитают жить в смешанных районах, в которых их число было бы лишь чуть больше, чем число «синих», а «синие», соответственно, тоже не предпочитают сегрегацию, но они не желают оставаться в меньшинстве среди «зеленых». Я выражу эти предпочте ния в виде цен, pg и pb, по которым «зеленые» и «синие», соответственно, хотели бы приобрести дом в данном районе, и пускай обе эти цены зависят от доли домов в районе, в которых проживают «зеленые», т. е. от f [0, 1]. Следующие уравнения показывают способ выражения вышеназванных предпочтений (2.1):
p |
(f) = 1/ |
(f + δ) – 1/ |
(f + δ)2 |
+ p, |
|
b |
2 |
2 |
|
|
|
p |
(f) = 1/ |
(f – δ) – 1/ |
(f – δ)2 |
+ p, |
(2.1) |
g |
2 |
2 |
|
|
|
где δ [0, 1/2], причем p — положительная константа, выражающая действи тельную стоимость идентичных домов. Дифференцируя обе функции по f и при равнивая результат к нулю, мы получаем, что идеальным районом для «зеленых» (для которого pg максимальна) является такой, в котором проживало бы 1/2 + δ «зеленых», в то время как «синие» предпочли бы такой, где «зеленых» 1/2 – δ. Поскольку разница между оптимальными районами (такими, за проживание в которых они заплатили бы наибольшие суммы) для «зеленых» и «синих» равна 2δ, я стану говорить о δ как о степени дискриминационных вкусов обоих типов людей (значение δ может различаться для двух групп, но может получиться и так, что одной из групп нет совершенно никакого дела до расового состава со седей). Пронормируем размер района к единице, чтобы говорить о числе «зеле ных» как о доле «зеленых».
Предположим теперь, что в каждом периоде некоторая часть «зеленых» и «синих», равная α, решает продать свои дома тем, кто живет за пределами райо на. Возможные покупатели, состоящие из людей двух типов в той же пропор ции, что и те, кто собирается продавать дома, посещают район. Доля возможных покупателей, относящихся к «зеленым», тогда равна f. Предполагаемые продав цы и покупатели выбирают друг друга случайным образом; предположим, что последние просто стучатся в дверь произвольно выбранного дома. Таким обра зом, в любой период ожидаемое число «зеленых», стремящихся продать свой дом и встречающихся с «синим» покупателем, равно αf (1 – f ). Каждый воз можный продавец за один период видит только одного покупателя и либо про дает дом, либо нет. Вероятность заключения сделки зависит от разницы между тем, во сколько оценивает дом покупатель и во сколько его оценивает продавец; обе цены задаются уравнениями (2.1). Таким образом, если «синий», желающий продать дом, встречает «зеленого», а значение f таково, что pg > pb, то сделка со вершится с вероятностью β (pg – pb), где β — положительная константа, показы вающая соотношение между разницей цен и вероятностью продажи.
Нам интересна эволюция распределения типов соседей с течением времени. Предполагая, что район достаточно велик, чтобы мы могли считать ожидаемые результаты приближенными к реальным значениям, и используя штрих (') для обозначения «следующего периода», мы можем записать f ′ как функцию от f, приняв во внимание, что в любом периоде некоторые «зеленые» вполне могут

Глава 2. Спонтанный порядок: самоорганизация экономической жизни 65
продать дом «синему», а некоторые «синие» — «зеленому». Таким образом, по лучаем
f ′ = f – αf (1 – f ) rb β(pb – pg) + α (1 – f ) f rg β (pg – pb ), |
(2.2) |
где rb = 1, если pb > pg, и равно нулю в остальных случаях, а rg = 1, если pg > pb , и равно нулю в остальных случаях (очевидно, что rb + rg = 1). Уравнение можно интерпретировать так: ожидаемая доля «зеленых» в следующем периоде равна их доле в данном периоде за вычетом тех, кто продаст дома «синим» (второй член выражения в правой стороне равенства), плюс «синие», что продадут свои дома «зеленым» (третий член выражения). Второй член выражения справа от знака равенства, например, показывает уменьшение количества «зеленых» изза продаж домов «синим»; αf — число «зеленых», ищущих покупателей, из них (1 – f ) встретятся с «синими», и если цена, предложенная «синим», окажется выше цены «зеленого», то продажа дома состоится с вероятностью β (pb – pg). Третий член выражения можно интерпретировать подобным же образом; если цена «зеленого» превысит цену «синего», то он продаст дом «зеленому». Зная, что pb + pg = 1, мы можем преобразовать уравнение следующим образом:
Df = f ′ – f = αf (1 – f ) β (pg – pb), |
(2.3) |
откуда ясно, что Df = 0, если pg = pb (т. е. сделок между потенциальными продав цами и покупателями различных типов не происходит, потому что покупатели не ценят дома выше, чем продавцы). Заметим, что Df = 0 также и в случае, когда f = 0 или f = 1 (т. е. когда осматривать дома приходят покупатели только того же типа, что и населяющие район однородные продавцы). Уравнение (2.3) называ ют «уравнением репликационной динамики». После преобразования его можно переписать в зачастую более удобном виде f = αf β( p g − p), где p — средняя цена, или p = fp g + (1 − f )pb .
Стационарное значение f является устойчивым равновесием, если экзоген ное изменение его значения в одну сторону приводит (за счет динамики, опи санной в уравнении (2.3.)) к появлению Df противоположного знака, т. е. если dDf/df < 0. Если неравенство выполняется, изменения f самокорректируются. Рисунок 2.1 иллюстрирует эту модель. Анализ рисунка (или незначительные вы числения) подтверждает, что «полузеленыйполусиний» район (Df = 0, потому что pg = pb) будет являться равновесием, однако неустойчивым (т. к. dDf/df > 0), поэтому случайное небольшое отклонение от такого распределения не приведет к самокоррекции, а станет накапливаться и приведет к совершенно сегрегиро ванному проживанию людей. Заметим также, что при δ < 1/4 как «зеленые», так и «синие» предпочтут совместное, а не сегрегированное проживание, даже если в результате такой сегрегации все их соседи окажутся одного типа с ними (это можно подтвердить, проверив, что pb (1/2) = pg (1/2) > pg(1) = pb (0)).
Таким образом, устойчивое равновесие с сегрегацией, которое, как мы и предполагали, становится единственным долгосрочным исходом этого взаимо действия, и оно хуже по Парето, чем ряд исходов со смешанным проживанием, которые нельзя реализовать как устойчивые равновесия в этой модели. Порази тельно, но результат сохраняется, даже если значение δ достаточно мало; полная
66 Часть I. Координация и конфликт: базовые социальные взаимодействия
Рис. 2.1. Спонтанная сегрегация жилищного сообщества. Две функции показывают максимальную готовность «синих» и «зеленых» платить за дом как функцию от f, «зеленой» доли сообщества. Отметим, что и «синие», и «зеленые» предпочитают смешанное соседство, проживая при этом в полностью сегрегированном сообществе соответствующего вида
сегрегация возникает, даже когда обе группы обладают фактически одинаковы ми вкусами, причем оптимальным для них районом был бы такой, в котором со отношение представителей обеих групп приближается к 50 : 50. Наконец, очень
легко показать, что полная сегрегация (любого типа) представляет собой устой чивое равновесие. Таким образом, районы станут локально однородными, в то
время как другие идентичные районы будут состоять только из людей другого типа, что в результате приведет к глобальной неоднородности. Каким именно станет состав района, исторически обусловлено: например, если в недавнем про
шлом значение f было меньше f *, нам стоит ожидать наступления случая f = 0. Причина провала координации, возникающего в таком случае, состоит в том, что, если домохозяйство принимает решение жить в неком обществе, его выбор влияет как на благополучие членов общества, к которому они присоединяются, так и того, из которого уезжают. Состав общества в нашем случае представляет собой некий «товар», который выбирает эта семья, и одновременно невольный продукт выборов всех семей. При этом не существует причины, по которой ре зультирующий исход стал бы эффективным, идет ли речь о том, что выбор проис ходит на основе предпочтения определенной расовой структуры (как это было здесь), высокого уровня образования у потенциальных соседей (Benabou, 1993), проживания среди собственников жилья (Hoff & Sen, 2002) или по другим при
знакам.
Япостроил модель процесса достижения рыночного равновесия, используя
вявном виде результаты социальных взаимодействий (кто с кем встречается, и что они делают). Индивиды пользовались только одним видом локальной ин

Глава 2. Спонтанный порядок: самоорганизация экономической жизни 67
формации: они не стремились найти наилучшую сделку, а просто производили трансакции с положительной вероятностью, пока это было взаимовыгодно, и никак иначе. Расовый состав соседей определился в процессе репликации, ко торый привел к заселению домов членами той или другой группы. Динамика изменения состава района была получена путем изучения того, какие места про живания реплицировали «цвет» своих хозяев, а какие его переменили. В гл. 6 я сопоставлю этот подход к социальному взаимодействию с моделированием рынков в вальрасовской модели.
моделирование эволюции поведения
Как расовый состав района, так и распределение норм индивидуального поведе ния или институциональных характеристик групп в популяции, а также их эво люция во времени зависят от того, какие признаки копируются, а от каких люди отказываются. «Признаки» — это любые характеристики индивида или группы, которые другие могут воспринять, отвергнуть или сохранить. Если дети като ликов склонны сохранять веру своих отцов, а дети протестантов — нет, то доля католиков в популяции вырастет (если предположить, что во всех семьях число детей одинаково и существуют только два типа религий). Если фирмы, признаю щие существование профсоюза своих работников, разоряются чаще, чем фирмы без профсоюзов, то новые фирмы начнут копировать фирмы более прибыльные, и профсоюзное движение проиграет.
Дифференциальная репликация может идти от людей и организаций, созна тельно стремящихся приобрести признаки, воспринять нормы или другие чер
ты, принесшие успех другим. Дифференциальная репликация, однако, может происходить и через процесс конформистского переноса, при котором репли
кация признаков является частотнозависимой, — к наиболее часто встречаю щимся признакам в популяции относятся более благосклонно1. Кроме того, хотя процесс дифференциальной репликации иногда носит спонтанный характер, он может быть насильственным, с применением силы со стороны государства, классов или организаций — как в случае, когда проигравшие войну вынуждены адаптироваться к культуре, системе обучения и законодательству победителей.
Детали процесса переноса очень важны, поэтому я рассмотрю эти и другие, более сложные, случаи в последующих главах, когда построю модель того, как эко номика и другие институты определяют формы эволюции предпочтений. Здесь
же я смоделирую важный, но чрезвычайно простой случай копирования успеш ного поведения. Процесс называется монотонной корректировкой платежей,
т. е. подразумевается класс механизмов переноса, обладающих тем свойством, что в них типы поведения с платежами выше среднего перенимаются другими
агентами; тем самым их доля в популяции увеличивается. Я также предположу, что пары взаимодействия между людьми образуются случайным образом.
1 Некоторые соображения по поводу важности конформистского переноса можно най ти в работах Бойда и Ричерсона (Boyd & Richerson, 1985) и Боулза (Bowles, 2001). Модель
конформистского обновления представлена в гл. 11.

68 Часть I. Координация и конфликт: базовые социальные взаимодействия
Предположим, что каждый член большой популяции обладает одним из двух признаков, исключающих друг друга (x и y)1. Этими признаками могут быть приверженность к определенным нормам поведения, гастрономические предпочтения или любые другие аспекты поведения, влияющие на выигрыши. Таким образом, x может означать «продавай товар по цене, равной предельным издержкам», «работай усердно», «заведи еще одного ребенка», «обменивайся подарками» или «ешь на завтрак здоровую пищу». Признак y станет обозначать противоположную норму для каждого случая. Эта модель легко расширяется
до варианта с большим количеством признаков. Я же рассматриваю модель с культурными признаками, приобретаемыми через обучение (у родителей или
других людей из предыдущего поколения, друзей и прочих), а не генетическое наследование. Таким образом, в данной модели корректировка поведения ста новится подобной переключению с одного признака на другой, а не наследова нию различий в признаке потомством (хотя эта модель легко адаптируется для случая с генетическим переносом признаков, что я покажу в примере с игрой «Ястреб — Голубь» чуть ниже). Меня интересует, сколько копий каждого при знака будет сделано в конце каждого периода (если индивид не оставил копий для следующего периода, следовательно, он переключился на другой признак; тот, кто оставил две копии, сохранил свой признак и скопировал его для другого). Заметим, что жизнь индивидов бесконечна, а индивиды являются просто пере носчиками признаков; именно о самих признаках можно говорить как о более или менее удачливых производителях своих копий. Размер популяции нормиро ван к единице.
Структура процесса переноса такова: индивиды приводят в действие страте гии, продиктованные им их признаками, в игре, в которой каждому, в зависимо сти от его поведения и поведения окружающих, приписываются некие платежи. Вслед за этим признаки реплицируются, причем те из них, чьи носители получи ли бо2льшие платежи, сделают относительно бо2льшее количество копий, меняя при этом частоту появления признаков в популяции. Предположим, что члены популяции случайным образом находят пару для того, чтобы взаимодействовать в симметричной игре с двумя агентами, платежи в которой мы обозначим как π (i, j) — платеж, который получит игрок, выбрав признак i в игре с партнером, имеющим признак j. Для любой доли признака x в популяции, т. е. для любого p [0, 1], ожидаемые платежи в таком случае равны
bx(p) = pπ (x, x) + (1 – p) π (x, y), |
|
by(p) = pπ (y, x) + (1 – p) π (y, y). |
(2.4) |
Прочтем первое уравнение: «С вероятностью p агент x находит себе партне ра x, и его платеж составляет π (x, x); а с вероятностью (1 – p) он находит себе партнера y, и тогда его платеж составляет π (x, y)».
1 Обзор математических моделей динамических систем, лежащих в основе представлен ных моделей, можно найти в работах Вейбулла (Weibull, 1995) или, более полно, в труде Хир ша и Смэйла (Hirsch & Smale, 1974).