

Регулировать экспрессию генов можно и после транскрипции
Основным объектом регулирующего влияния на синтез белка и нуклеиновых кислот является транскрипция. Однако существует возможность регуляции и после транскрипции. Это происходит на всех этапах процессинга РНК, трансляции и посттрансляционной модификации белковых молекул.
Аттенуация транскрипции один из возможных механизмов регуляции экспрессии генов.
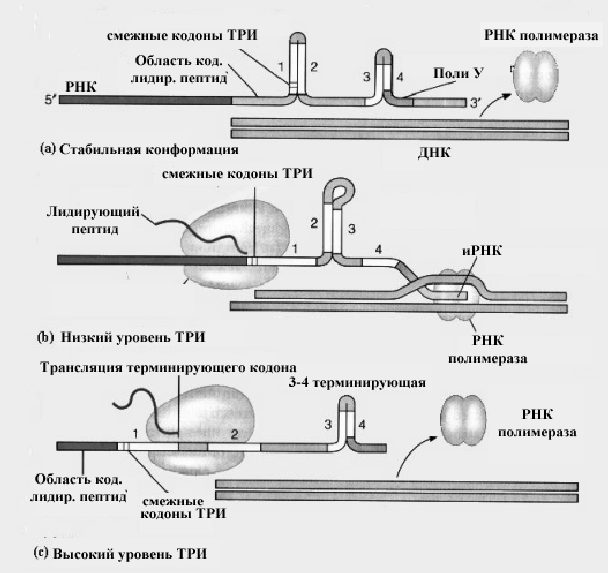
У прокариот существует возможность остановить начавшуюся транскрипцию, благодаря специальной последовательности нуклеотидов на РНК, которая обеспечивает удаление РНК-полимеразы с матрицы ДНК. Это хорошо показано на примере упоминаемого выше триптофанового оперона кишечной палочки. У оперона триптофана имеется вторая регуляторная область, область аттенуации (ослабления).
 Этот
участок, расположен приблизительно на
140 нуклеотидов вниз к 3’концу от участка
инициации иРНК. Синтез иРНК может быть
преждевременно остановлен на этом
участке, если уровень триптофана
повышается выше некоторого значения.
Механизм ослабления показан на рис .
Лидирующий транскрипт кодирует
14-амино-кислотный олигопептид, который
содержит два смежных остатка триптофана.
Он также содержит четыре облас
Этот
участок, расположен приблизительно на
140 нуклеотидов вниз к 3’концу от участка
инициации иРНК. Синтез иРНК может быть
преждевременно остановлен на этом
участке, если уровень триптофана
повышается выше некоторого значения.
Механизм ослабления показан на рис .
Лидирующий транскрипт кодирует
14-амино-кислотный олигопептид, который
содержит два смежных остатка триптофана.
Он также содержит четыре облас
Рис.Аттенуация транскрипции на примере триптофанового оперона Е.coli.
ти (1, 2, 3, и 4) способные к формированию трех шпилек, SL-1:2, SL-2:3, и SL-3:4. SL-3:4 - участок терминации транскрипции; он непосредственно следует за отрезком поли У. Шпилька SL-3:4 может образоваться, если предотвратить образование шпильки SL-2:3.
В отсутствии триптофана (то есть, когда необходима полная экспрессия оперона), рибосома задерживает два кодона триптофана, расположенные в области 1. В результате, SL-1:2 не может формироваться, а вместо этого формируется шпилька SL-2:3, что в свою очередь предотвращает формирование шпильки SL-3:4, и синтез иРНК не прерывается, так как SL-2:3 не узнается как сигнал терминации транскрипции.
В присутствии триптофана, однако, рибосома переместится в область 2 лидирующего транскрипта прежде, чем та образует пары оснований с областью 3, таким образом, не допуская формирования SL:-2:3. Это способствует образованию SL-3:4, что приводит к преждевременной терминации транскрипции. Аттенуация транскрипции происходит и в случае, когда останавливается
Следует заметить, что аттенуация транскрипции, по-видимому, ведущий способ регуляции и других оперонов, привлекаемых для биосинтеза аминокислот. Лидирующие последовательности иРНК, образующиеся при транскрипции оперонов гистидина, треонина и фенилаланина содержат по 7 или 8 кодонов для соответствующей аминокислоты. Это делает аттенуацию транскрипции удобной формой регуляции активности оперонов в зависимости от уровня аминокислот в цитоплазме.
П о
понятным причинам такой механизм не
пригоден для клеток эукариот, однако
аттенуация используется и в эукариотических
клетках, но роль белков, контролируюющих
прохождение РНК-полимеразы через участки
аттенуации берут на себя белковые
ансамбли промотора.
о
понятным причинам такой механизм не
пригоден для клеток эукариот, однако
аттенуация используется и в эукариотических
клетках, но роль белков, контролируюющих
прохождение РНК-полимеразы через участки
аттенуации берут на себя белковые
ансамбли промотора.
Рис.. Позитивный и негативный контроль сплайсинга в экспрессии одинаковых генов разными клетками
Альтернативный сплайсинг рнк -своеобразная форма регуляции экспрессии генов
На рис. показаны возможные варианты позитивного (2) и негативного (1) контроля сплайсинга в двух типах клеток. Молекула репрессора тормозит сплайсинг первичного транскрипта РНК в клетке 2, а молекула активатора включает механизм сплайсинга в той же клетке. Это позволяет создавать различные белки с первичных транскриптов одного и того же гена в разных клетках или переключать образование функционально активных и неактивных белков в пределах одной клетки.
Присоединение поли А к 3’ концу РНК может контролироваться клеткой.
В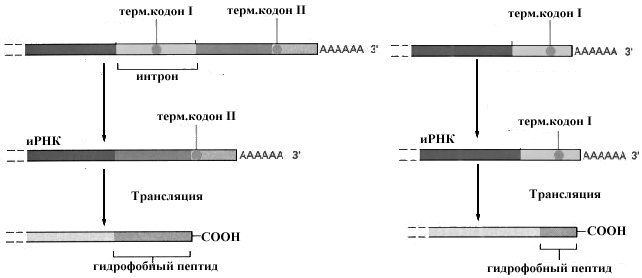 отличие от бактерий, 3’ конец первичного
транскрипта РНК у эукариот подвергается
полиаденилированию после предварительного
удаления отрезка нуклеотидов. В некоторых
случаях клетка может контролировать
место расщепления, обеспечивая образование
молекул иРНК различной длины, что
оказывает влияние на структуру белковой
молекулы. Хорошо изученным примером
такой формы регуляции является синтез
мембраносвязанных и растворимых форм
антител у активированных антигеном и
неактивированных В лимфоци
отличие от бактерий, 3’ конец первичного
транскрипта РНК у эукариот подвергается
полиаденилированию после предварительного
удаления отрезка нуклеотидов. В некоторых
случаях клетка может контролировать
место расщепления, обеспечивая образование
молекул иРНК различной длины, что
оказывает влияние на структуру белковой
молекулы. Хорошо изученным примером
такой формы регуляции является синтез
мембраносвязанных и растворимых форм
антител у активированных антигеном и
неактивированных В лимфоци
Рис.10-27.Схема регуляции синтеза мембраносвязанных и растворимых форм антител.
тов. Секретируемые молекулы антител идентичны мембраносвязанным за исключением строения С-концевого отдела молекулы, который у мембраносвязанных антител представлен длинным отрезком гидрофобных аминокислот, обеспечивающих встраивание этих молекул в мембрану. Это объясняется тем, что первичные транскрипты активированного и неактивированного антигеном лимфоцита различаются по длине. Короткий транскрипт активированного лимфоцита не содержит участка, кодирующего гидрофобные аминокислоты и образующиеся после трансляции белки не задерживаются в мембране, а секретируются из клетки.