

10.3. Биосинтез терпеноидов
Еще один анаболический путь превращения ацетил-СоА представлен био-синтезом терпеноидов. Он начинается с образования ацетоацетил-СоА по схеме сложноэфирной конденсации, катализируемой тиолазой:
![]()
Еще одна молекула ацетил-СоА реагирует с ацетоацетил-СоА по схеме альдольной конденсации с образованием 3-гидрокси-3-метилглутарил-СоА при катализе соответствующей синтазой:

Теперь гидроксиметилглутарил-СоА-редуктаза переводит этот тиоэфир ди-карбоновой кислоты в диоксикарбоновую кислоту, называемую мевалоно-вой кислотой:

Соответствующие киназы переносят на гидроксильные группы этого ве-щества один фосфатный и один пирофосфатный фрагмент от трех молекул АТФ:

Эта фосфорилированная мевалоновая кислота отщепляет фосфат и декарб-оксилируется с образованием изопентенилпирофосфата:
 ,
,
который по схеме гидрирования-дегидрирования может превращаться в ди-метилаллилпирофосфат. Далее идет последовательное алкилирование изо-пентенилпирофосфата диметилаллилпирофосфатом с образованием гера-нилпирофосфата и далее акилирование изопентенилпирофосфата геранил-пирофосфатом с образованием фарнезилпирофосфата:


При биосинтезе стероидов две молекулы фарнезилпирофосфата соединя-ются по атомам углерода, несущим пирофосфатные остатки (промежу-точным продуктом является пресквален-пирофосфат с циклопропановым фрагментом), и образуется сквален:

Сквален окисляется скваленэпоксидазой с образованием 2,3-эпоксида, ко-торый в активном центре скваленоксидциклазы сворачивается в структу-ру, обеспечивающую образование тетрациклического ланостерина в ре-зультате протонирования, раскрытия эпоксидного цикла и следующих за этим атак карбокатионов по двойным связям:
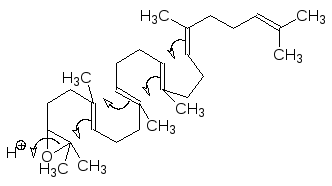
Образовавшийся в результате циклизации карбокатион стабилизируется, отщепляя протон, и превращается в ланостерин с одновременной мигра-цией двух метильных групп:

Далее идут окислительные превращения с отщеплением метильных групп, с гидрированием двойных связей, дегидрированием с образованием двой-ных связей и другие реакции, приводящие к образованию холестерина, эргостерина и других стероидных липидов, витамина D, стероидных гор-монов, желчных кислот.
Изопентенилпирофосфат является также базовым веществом для образо-вания витаминов А, Е и К, каротиноидов, каучука и других полиизопренов, фитольной компоненты хлорофилла и других полипренольных остатков, например, в убихиноне.
10.4. Биосинтез аминокислот
Растения и микроорганизмы синтезируют все белковые (протеиногенные) аминокислоты и большое число небелковых аминокислот, участвующих в биосинтезе самых разных биогенных соединений, включая, антибиотики, токсины и другие защитные молекулы. При изучении метаболизма уже рассматривалось образование аминокислот из -кетокислот в результате действия трансаминаз, поэтому биосинтез аланина из пирувата, глютами-новой кислоты, глютамина, аспарагиновой кислоты и аспарагина из кето-глутарата и, соответственно, оксалоацетата уже был представлен выше. Достаточно простая схема реализуется при образовании пролина из ‑полу-альдегида глютаминовой кислоты с его циклизацией в карбоксизамещен-ный пирролин и восстановлением С=N-связи:

Так же легко прослеживается путь от глицеринового альдегида к серину, а глицин образуется из серина в результате отщепления концевой гидрокси-метильной группы в виде формильного фрагмента под действием тетра-гидрофолат-зависимого фермента.
В растениях из серина образуется также цистеин в результате алкили-рования сероводорода О-ацетилсерином в присутствии цистеин-синтазы:

В свою очередь цистеин в растениях служит источником серы для образо-вания метионина, а у животных источником серы служит уже эта незаме-нимая аминокислота. Для получения сбалансированных по аминокислот-ному составу кормов метионин производят в промышленном масштабе по схеме:


Окислением пропилена кислородом воздуха на оксиде ванадия в качестве катализатора получают акролеин, который присоединяет метилмеркаптан по олефиновой двойной связи с образованием пропионового альдегида с метилтиольной группой в -положении. Это вещество вступает в реакцию Бухерера-Бергса и превращается в замещенный гидантоин, который гидро-лизуется в присутствии щелочи при нагревании под давлением и превра-щается в D,L-метионин. Его способом Гринштейна (ацетилирование по аминогруппе и ферментативный гидролиз ацетилазой) можно разделить на стереоизомеры, но для метионина эта операция не является обязательной, поскольку его D-изомер нетоксичен.
Рассмотрение синтетических путей, ведущих ко всему разнообразию про-теиногенных и небелковых аминокислот, можно ограничить схематичес-ким представлением образования в растениях фенилаланина и аминокис-лот с алифатическими заместителями (валина, лейцина, изолейцина), по-скольку на блокировке их биосинтеза основано фитотоксическое действие самых распространенных гербицидов.
Образование всех ароматических соединений в растениях начинается с шикимовой кислоты, образующейся в ряде последовательных реакций из D-эритрозо-4-фосфата и фосфоенолпирувата. 5-Фосфошикимовая кислота реагирует с фосфоенолпируватом при катализе кислотой с образованием 3‑еноилпирувил-5-фосфошикимовой кислоты:

Образовавшееся соединение отщепляет фосфат и превращается в хоризмо-вую кислоту:

В результате внутримолекулярного алкилирования хоризмовая кислота превращается в префеновую кислоту, которая после декарбоксилирования и дегидратации превращается в фенилпировиноградную кислоту:

Декарбоксилирование и дегидрирование префеновой кислоты приводит к 4-гидроксифенилпировиноградной кислоте, которая действием трансами-наз превращается в тирозин, а фенилпировиноградная кислота превраща-ется в фенилаланин. Из хоризмовой кислоты через стадию образования антраниловой кислоты в клетках растений образуется триптофан.
Взаимодействие фосфошикимовой кислоты с фосфоенолпируватом – это одна из немногих биохимических реакций, когда фосфоенолпируват при-нимает участие в протонированном виде. Предполагалось, что исполь-зуемый в качестве средства борьбы с сорной растительностью фосфономе-тилглицин является антиметаболитом протонированного фосфоенолпиру-вата:
 и
и

Однако, более детальные исследования показали, что сайты связывания этих веществ различны, то есть речь идет о неконкурентном ингибирова-нии 3-еноилпирувил-5-фосфошикимат-синтазы фосфонометилглицином. Обработка растений этим веществом приводит к их гибели в течение нес-кольких часов из-за нарушения биосинтеза ароматических кислот.
Биосинтез валина основан на реакции алкилирования пирувата по карбо-нильному атому углерода гидроксиэтильным остатком, образующимся при декарбоксилировании другой молекулы пирувата на тиаминпирофосфате в активном центре ацетолактатсинтазы:

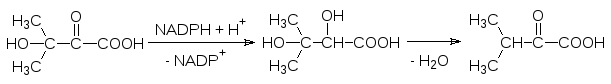
Затем молекула ацетолактата претерпевает внутримолекулярную пере-группировку: после протонирования атома кислорода карбонильной груп-пы происходит миграция метильной группы в виде карбаниона на карбо-нильный атом углерода, а протон отходит от гидроксильной группы. Про-дуктом этой реакции становится 2‑кето-3-гидроксиизовалериановая кислота:

Образовавшаяся гидроксикетокислота в результате восстановления NADPH-зависимым ферментом превращается в 2,3-дигидроксиизовалериа-новую кислоту, которая дегидратируется и превращается в -кетоизовале-риановую кислоту. Трансаминаза переводит ее валин:
-Кетоизовалериановая кислота является также исходным продуктом для биосинтеза лейцина. По аналогии с образованием валина, но с -кетобути-ратом, образующимся из треонина, идет биосинтез изолейцина,.
Ключевой фермент в биосинтезе алифатических аминокислот – ацетолак-татсинтаза – блокируется сульфонилгетерилмочевинами, например хлор-сульфуроном (Глин):

Этот гербицид, относящийся к третьему поколению агрохимических пре-паратов, имеет нормы расхода около 5 г/га. Получено также большое чис-ло его структурных аналогов, которые более избирательны и более легко подвергаются метаболической деградации в растениях и в почве.