

Лекции по экологической физиологии / Лекция 2 ЭФ 2012
.doc
Лекция 2
Энерготраты в условиях двигательного покоя
Метаболизм покоя и структура тела животных
Энерготраты в условиях двигательного покоя
Базальный метаболизм в обычной жизни, в лучшем случае, протекает во сне. Самые мелкие теплокровных, у которых наиболее высокая скорость окислительных процессов в тканях и наибольшая потребность в кислороде, спят только короткими промежутками времени, а значительную часть суток двигаются в поисках пищи. При более длительном сне у таких теплокровных снижается температура тела. Это происходит с мелкими птичками калибри ночью, когда они не могут летать из-за плохого зрения, или с летучими мышами днем, которые ночью ловят насекомых, а днем спят в своих прохладных помещениях. Поэтому очень важно оценить затраты энергии теплокровного не только в состоянии его БМ, но и в условиях повседневной жизни. Методически сделать это трудно, а порой и невозможно, так как трудно организовать длительные круглосуточные наблюдения животных, причем в обстановке, приближенной к их обычной среде обитания.
Впервые такие исследования на млекопитающих провели в 30-40-х годах прошлого столетия А.Д. Слоним с коллегами. Эти опыты были уникальны, так как впервые в них использовались дикие животные. Для измерения скорости потребления О2 животными в условиях естественного покоя (назовем это состояние «метаболизм в покое ПМ») их натощак помещали в сравнительно просторные респираторные камеры, к которым предварительно приучали и в которых Т воздуха поддерживалась на 5оС ниже их Т комфорта (при Т комфорта измеряется БМ). Результаты этих опытов нашли отражение в монографии, вышедшей в 1952 г «Животная теплота и ее регуляция в организме млекопитающих» [46]. Приведу два рисунка из этой книги, рис. 6.
|
|
|
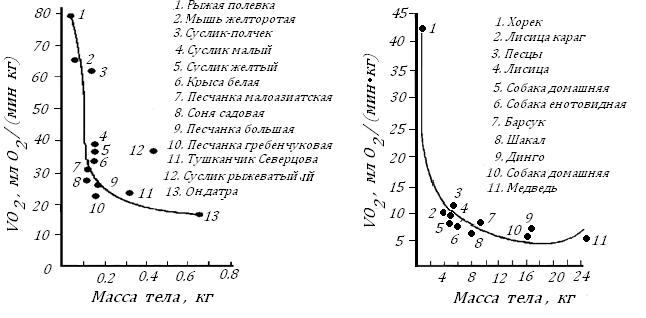
Рис. 6. Скорость потребления О2 грызунами (слева) и хищниками (справа) в условиях двигательного покоя натощак при Т воздуха на 5ОС ниже Т комфорта [46].
Первый рисунок показывает, что VО2/кг у грызунов по мере снижения Мт вначале меняется незначительно, но резкое растет, когда Мт падает от 300 г до 30 г. Похожая картина наблюдается у хищников. Их кислородная потребность мало меняется до Мт в 6 кг и резко увеличивается при снижении ее до 50 г. Эти исследования показали, что в привычной среде обитания с Т более низкой, чем Т, при которой измеряют БМ, энергетические затраты резко растут у наиболее мелких теплокровных. Ниже сравним величины VО2, измеренные в условиях двигательного покоя (ПМ согласно рис. 6), с расчетным БМ согласно формуле БМ=10 Мт-0.25 у грызунов и хищников с малой (слева) и большой (справа) Мт:
|
Мт, г |
мл 02мин-1кг-1 |
ПМ/БМ |
||
|
БМ |
ПМ |
|||
|
Грызуны Хищники |
30 и 300 50 и 6000 |
24 и 14 21 и 6.4 |
78 и 22 41 и 9.4 |
3.2 и 1.6 2.0 и 1.5 |
Видим, чем мельче теплокровное, тем больше ПМ превышает у него БМ. Так, у маленьких грызунов и хищников соотношение ПМ/БМ равняется 3.2 и 2.0, а у крупных животных оно падает до 1.6 и 1.5.
Данные на рис. 6 иллюстрируют и видовое своеобразие этого показателя: он повышен у собаки динго по сравнению с более мелким шакалом. Замечено, что у тропических и субтропических видов ВМ и ПМ ниже, чем у близких к ним видах теплокровных, но живущих в северных зонах [46, 53].
Самое интересное, что зависимость минимального окислительного обмена от поверхности тела, правильнее сказать, от Мт0.75, характерна не только для теплокровных. Этот закон работает для всего живого, даже для растений [71].
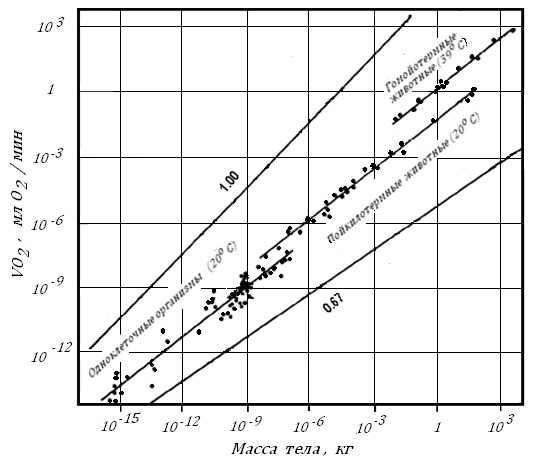
Рис. 7. Скорость энергетических затрат живых организмов в зависимости от их массы тела в состоянии, близком к базальному метаболизму, (Hemmingsen, 1960), по: [71].
На рис. 7 приведены аллометрические зависимости для одноклеточных с Мт от 10-6 мкг до 1 мкг, холоднокровных от одного до 105 г и у теплокровных, о которых говорилось ранее. Данные о VО2 для пойкилотермных организмов, приведенные на этом рисунке, получены при произвольно выбранной, но одинаковой для всех животных Т среды; в этих условиях состояние окислительного метаболизма у них , по-видимому, приближалось к ПМ. График показывает, что средняя потребность в кислороде животных всех групп была пропорциональна Мт в степени 0.75.
Недавно группа ученых провела оценку затрат энергии на жизнедеятельность у 3006 видов животных и растений в условияхих относительного покоя при обычной для гомойотермных Т тела и при 25ОС у пойкилотермных. В этой работе были использованы многочисленные опубликованные ранее данные, рис. 8.
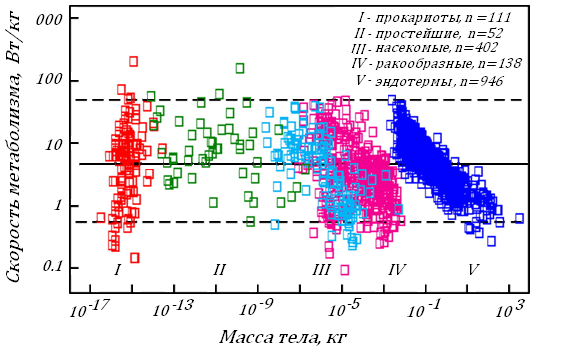
Рис. 8. Энергетические затраты живых существ в зависимости от их массы тела [95]. Сплошная линия показывает среднюю величину этих затрат, 4.7 вт/кг или 14 мл О2/(мин∙кг); 95% видов, принадлежащих к пяти группам существ, располагаются между верхней и нижней прерывистых линий (49 и 0.51 вт/кг). Т тела для пойкилотермных организмов 25ОС.
Оказалось, что величина энергетических затрат, отнесенная к 1 кг Мт, у всех живых существ, несмотря на 1020-кратную разницу их в Мт, находится в довольно узких пределах: 0.5-50 вт/кг, а для 95% организмов - всего в 30-кратных пределах 0.3-9 вт/кг или 0.9-27 мл О2мин-1кг-1.
Исследователи называют этот предел «оптимальный» для живых существ и связывают его с биохимической универсальностью живых структур. Это означает, что зависимость скорости окислительного метаболизма в целом для живого мира не зависит от Мт в степени 0.75 или 0.67; она определяется исторически сформированными и общими для всех живых существ скоростями основополагающих клеточных реакций, рис. 8.
Однако и в этом «оптимальном» энергетическом пределе сохраняется зависимость энергетических трат от Мт, и ее можно видеть на рис. 8 для всех групп организмов, кроме прокариот. Особенно заметна она у теплокровных.
Метаболизм покоя и структура тела животных
С помощью каких механизмов осуществляется связь массы и поверхности тела животного с уровнем его БМ? Какие изменения процессов и структур в организме приводят по мере увеличения Мт к снижению потребности его тканей в кислороде (в расчете на 1 кг Мт)? При этом у теплокровных такая связь обеспечивает постоянство их температуры тела.
Оказывается, если выстроить зрелых животных, например, одного класса, в ряд по мере увеличения их Мт (рис. 4), то многие структурные и функциональные параметры их организма будут закономерно меняться. Так, с увеличением размера теплокровного, как правило, уменьшается относительная масса внутренних органов – основных потребителей кислорода, но сохраняется неизменными относительная масса скелетных мышц и несколько увеличивается масса покровных и опорных структур. Приведу примеры таких изменений у млекопитающих.
Таблица 7. Относительная масса органов, % Мт (кг),
у млекопитающих разного размера, [73, 101, 102]
|
Легкое |
1.13Мт–0.01 |
|
Сердце |
0.58Мт–0.02 |
|
Кровь |
6.2Мт–0.02 |
|
Почки |
0.73Мт–0.1 |
|
Печень |
3.33Мт–0.13 |
|
Головной мозг |
1.0Мт–0.3 |
|
Скелетные мышцы |
~40Мт00 |
Табл. 7 показывает, что показатель степени при Мт для всех внутренних органов и головного мозга отрицательный. Это означает, что по мере увеличения Мт относительная масса этих органов снижается; сохраняется лишь масса скелетной мускулатуры (показатель степени 0). Она, в среднем, близка к 40% Мт. Однако у животных с низкой и высокой двигательной активностью масса скелетных мышц различается, находясь в пределах 25-45% [90].
У самых мелких теплокровных масса скелетной мускулатуры, как правило, несколько ниже, а масса внутренних органов выше. В целом, в ряду теплокровных по мере увеличения Мт относительная масса легких, сердца, почек, печени и, особенно, головного мозга снижается; эти органы обладают высоким кислородным запросом, что приводит и к снижению БМ.
Поэтому не случайно наиболее резкое снижение БМ и ПМ в связи с падением Мт происходит в группе мелких теплокровных (табл. 4). У водных млекопитающих, проводящих основную часть жизни в сравнительно холодной воде и имеющих значительный слой подкожного жира (до 60% Мт), массы внутренних органов и скелетных мышц, соответственно ниже, приводимых в табл. 7.
С увеличением размера животного, особенно наземного, требуется больше тканей, сохраняющих и поддерживающих форму всего тела и его органов.
Таблица 8. Структурные параметры сердца у млекопитающих
разного размера (Black-Shaffer et al., 1965), по: [75]
|
Вид |
Масса сердца, кг |
Диаметр волокна, мкм |
Толщина стенки желудочка, мм |
|
|
мышца |
соединительная ткань |
|||
|
Девочка 15 лет |
0.18 |
13 |
10 |
0 (0%) |
|
Слон индийский |
7.3 |
18 |
40 |
0.23 (0.6%) |
|
Кит синий |
290 |
17 |
140 |
1.5 (1.1%) |
Поэтому в его покровных и опорно-двигательных структурах увеличивается доля соединительной ткани, что неизбежно снижает долю мышечных структур. В табл. 8 показано, как с увеличением размера сердца у млекопитающих в стенке его желудочков растет толщина соединительной ткани.
Передвигаясь по ряду млекопитающих в сторону крупных видов, можно видеть, что у них не только снижается относительная масса внутренних органов, но падает интенсивность их работы. Например, частота сокращения сердца и дыхательных мышц снижается. Замечено интересное явление: число сокращений сердца в течении всей жизни значительно не различается, например, у млекопитающих с очень разной Мт. Поэтому чем выше базальный метаболизм, тем чаще пульс и короче жизнь, табл. 9.
Таблица 9. Аллометрическая зависимость некоторых функциональных параметров от массы тела (кг)
млекопитающих [101, 102]
-
Частота дыхания в минуту
53.5Мт–0.26
Частота сердцебиений в минуту
241Мт–0.25
Время жизни, годы
7.52Мт0.29
Число сердцебиений за жизнь
9.5 108Мт0.04
На общей схеме сердечно-сосудистой системы (ССС, рис. 9) можно видеть путь, который проходит кровь у разных позвоночных, перенося кислород из внешней среды (воды или воздуха) к тканевым клеткам.
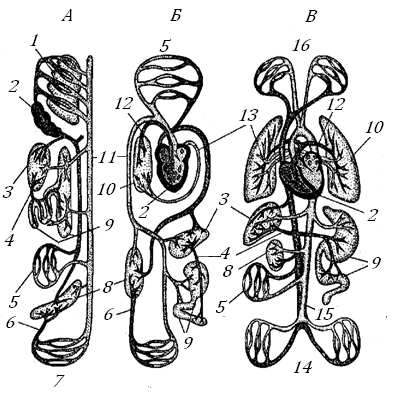
Рис. 9. Три типа кровеносной системы позвоночных, показывающих развитие легочных (или жаберных) сосудов, сосудов большого круга и воротных систем почек и печени [13]
А – рыба, Б – амфибия, В – млекопитающее. 1 – жабры, 2 – сердце, 3 – печень, 4 – воротная вена печени, 5 – ткани тела, 6 – воротная вена почек, 7 – сосуды к хвосту иот него, 8 – почки, 9 – пищеварительный тракт, 10 – легкие, 11 – спинная аорта, 12 – легочная артерия, 13 – легочная вена, 14 – сосуды к конечностям и от них, 15 – аорта, 16 – сосуды к голове и передним конечностям и от них.
Одной из главных характеристик работы сердечно-сосудистой системы является минутный объем кровообращения МОК - количество крови, которое каждый желудочек выбрасывает за 1 минуту в малый и большой круги кровообращения.
На рис. 10 можно видеть, что в ряду теплокровных МОК. растет медленнее, чем увеличивается их Мт. В покое величина МОК у млекопитающих пропорциональна Мт, примерно, в степени 0.75.
Заметим, что МОК не удается измерить у самых крупных и самых мелких животных. Поэтому у этих животных его величину можно рассчитать, умножив частоту пульса на диастолический объем желудочков, измеряемый посмертный или рассчитанный по их массе.
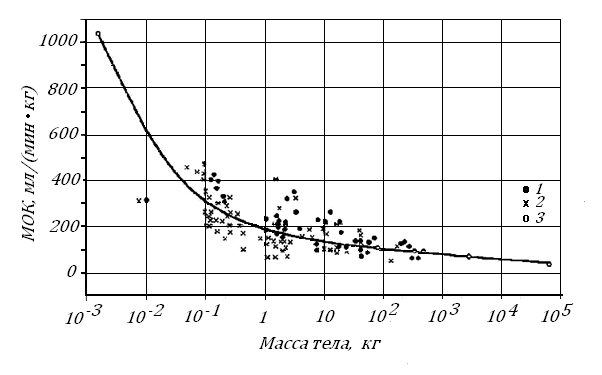
Рис.10. Зависимость минутного объема кровообращения у млекопитающих от их массы тела в состоянии двигательного покоя [74, 75]. 1, 2, 3 – животные: без наркоза, в наркозе, их состояние не указано или расчетные данные.
Похожая зависимость МОК от Мт характерна и для других теплокровных – птиц: на 9 видах птиц от волнистого попугайчика до страуса Эму (Мт в этом ряду росла от 35 г до 38 кг) МОК в условиях двигательного покоя менялся согласно уравнению МОК (мл/мин) = 291Мт0.69(кг), по [89].
Если в ряду теплокровных Мт растет быстрее, чем увеличивается объем сердечного выброса, то кровоток, рассчитанный на единицу Мт, будет падать. Так, у человека с Мт 70 кг по сравнению с мышью (Мт 2 г) удельный МОК снижается, примерно, в 10 раз: 1000 и 100 млмин-1кг-1. Это означает, что объемная скорость кровотока (ОСК) в органах должна падать. Однако это падение не одинаково в различных органах.
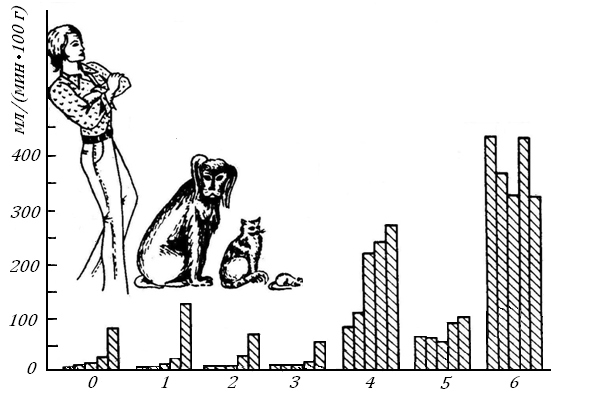
Рис. 11. Объемная скорость кровотока в органах млекопитающих с массой тела 70, 20, 2, 0.2 и 0.02 кг [74, 75]. Расположение столбиков (слева направо) показывает величину органного кровотока, соответствующую пяти видам млекопитающих от человека до мыши : 0-МОК, 1-кожа, 2-скелет, 3-скелетные мышцы, 4-сердце, 5- органы v. porta, 6-почки.
Рис. 11 показывает: чем крупнее животное, тем больше снижается кровоток в его опорно-двигательных структурах – коже, костях и скелетной мускулатуре. В сердце и желудочно-кишечном тракте с печенью (органы, из которых кровь вытекает по печеночной вене) он снижается не столь значительно и меньше, чем снижается сам МОК/кг Мт; в почках ОСК, возможно, остается независимой от Мт и наиболее высокой.
Это не удивительно: функция почки - выделение воды и излишка солей из организма – требует очень высокого почечного кровотока, и только малая часть его используется для переноса кислорода в почку. Для других органов, представленных на рис. 11, очевидно: чем выше интенсивность окислительного метаболизма в органных клетках, тем больше требуется им кислорода и тем больший поток крови должен протекает через орган.
Кислород поступает в организм через дыхательные органы; там же выделяется во внешнюю среду углекислый газ, образующийся в клетках. В дыхательных органах располагается первый барьер, который встречает кислород на пути из внешней среды к органным клеткам. У насекомых этот барьер наиболее прост, кислород в их ткани поступает через узкие воздушные трубки – трахеи. У водных позвоночных органами дыхания служат жабры и кожа с сетью кровеносных капилляров, через стенку которых происходит газообмен. У всех теплокровных, водных и наземных, имеются легкие, и первым барьером для кислорода служат структуры, отделяющие воздух в легочных альвеолах от крови в легочных капиллярах, рис. 12.
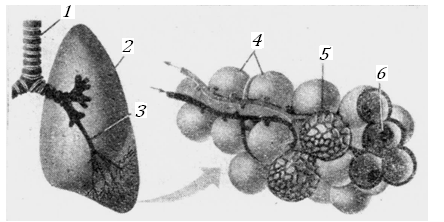
Рис. 12. Схематическое строение легких теплокровных.
1, 2, 3 – трахея, легкое, бронх; 4, 5. 6 – альвеолы, сеть капилляров, аьвеолярные ходы [13].
Возникает естественный вопрос, меняются ли органы внешнего дыхания у животных по мере увеличения их Мт. Оказывается объем легких, а большую часть его занимают альвеолы, у млекопитающих растет пропорционально их Мт1.06, рис. 13.
Увеличение альвеолярной поверхности несколько отстает от увеличения объема легких, будучи пропорциональным ему в степени 0.89, рис. 14.
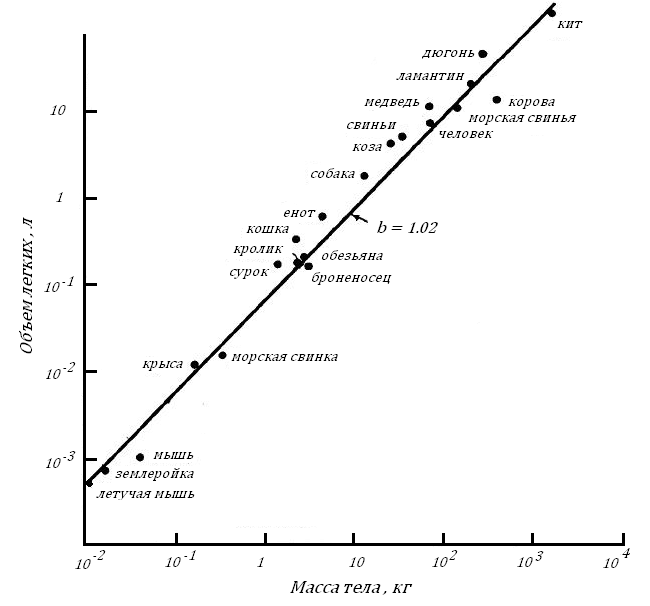
Рис. 13. Зависимость объема легких от массы тела у млекопитающих (Tenney, Remmer, 1963), по: [73].
Оба графика показывают, что объем воздуха, с которым контактирует кровь в легочных капиллярах, и величина альвеолярной поверхности увеличиваются по мере увеличения Мт животных больше, чем растет их внешняя поверхность тела и больше, чем растет их БМ (эти показатели увеличиваются пропорционально Мт в степени 0.67 и 0.75, соответственно).
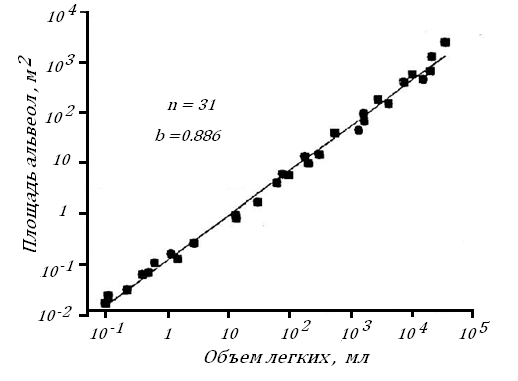
Рис. 14. Связь альвеолярной поверхности легких с его объемом у млекопитающих c разной массой тела [100].
Транспорт дыхательных газов между тканевыми клетками и внешней средой (для теплокровных это - альвеолярный воздух) производится током крови: из тканевых капилляров, расположенных в большом круге кровообращения, кровь выносит избыток СО2, а из легочных капилляров в малом круге она переносит к тканям кислород.
Кислород транспортируется в гемоглобине (Hb) – белковой молекуле с четырьмя протопорфиринами, в каждом из которых имеется двухвалентный ион железа (они образуют пигмент гем). М.м. гемоглобина 64500. Каждая его молекула может присоединить 4 молекулы кислорода, образуя оксигемоглобин (HbO2). Средняя концентрация Hb в крови здоровых мужчины 158 г/л, женщины 140 г/л. 1 г Hb может присоединить 1.36 мл О2.
Количество кислорода, превращающее Hb в HbO2, зависит от концентрации кислорода в окружающем растворе или в плазме крови, если речь идет о Hb в эритроцитах. Эту зависимость изучают в специальной ячейке, в которой сохраняют постоянным парциальное давление СО2 и температуру и снижают парциальное давление О2 (рО2). В ячейку помещают насыщенную кислородом кровь и определяют, как меняется в ней концентрация HbO2.
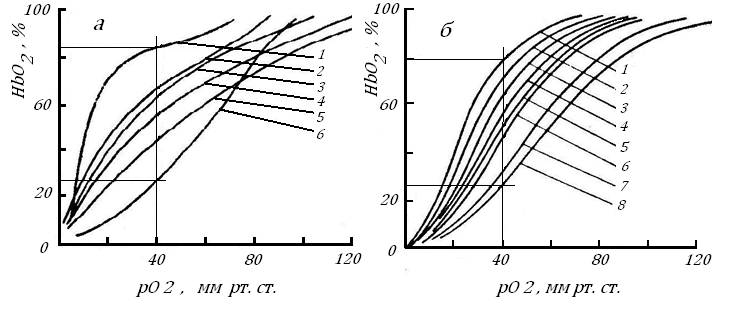
Рис. 15. Кривые диссоциации оксигемоглобина у млекопитающих с разной массой тела, а - (Филатова, 1944), по [47], б - [73].
По оси абсцисс – рО2 в среде, по оси ординат - концентрация HbO2, %. Звери на левом рисунке: 1 - лошадь, 2 - овца, 3 - степной хорек, 4 - кошка, 5 - желтый суслик, 6 - морская свинка; на правом рисунке: 1 - слон, 2 - лошадь, 3 - человек, 4 - овца, 5 - лиса, 6 - кошка, 7 - крыса, 8 – мышь
Кривая диссоциации HbO2 имеет S-образную форму: по мере снижения рО2 в растворе, окружающем эритроциты: в начале насыщенность гемоглобина кислородом не меняется, оставаясь максимальной, затем происходит его резкое снижение и, когда рО2 в среде падает до нуля, весь кислород покидает молекулу HbO2.. На рис. 15 можно видеть: чем мельче млекопитающее, тем слабее связь кислорода с молекулой Hb. При рО2, равном 40 мм рт. ст. (обычное рО2 для венозной крови у этих животных), концентрация кислорода в крови у мелких видов в 2 и более раз ниже, чем у крупных. Это означает, что чем мельче теплокровное и выше у него скорость окислительных реакций, тем больше кислорода покидает его кровеносное русло.
Высокая плотность капилляров в тканях мелких теплокровных способствует большему выходу кислорода из крови в окружающие капилляр структуры. По мере увеличения Мт их плотность падает, что хорошо видно на примере миокарда, рис. 16.
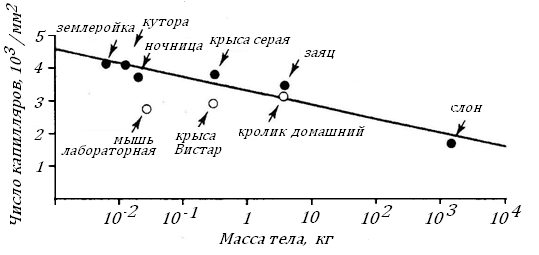
Рис. 16. Плотность кровеносных капилляров на поперечных срезах желудочка сердца у диких и домашних видов млекопитающих с разной массой тела [99].
Кроме того, по мере увеличения размера тела теплокровного скорость потребления кислорода его органными клетками снижается. Такой процесс наблюдается в клетках печени в ряду птиц с Мт от 13 г (ткачик) до 35 кг (страус эму). Дополним, что снижение VО2 в гепатоцитах присходит одновременно с уменьшением относительной массы печени, рис. 17.
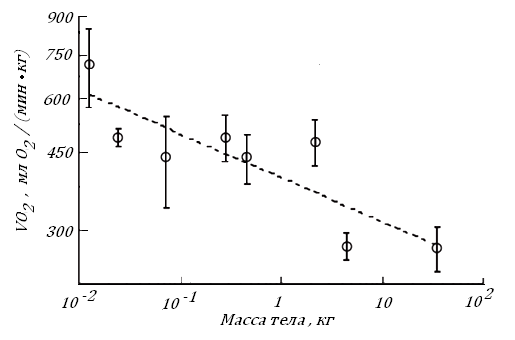
Рис. 17. Скорость дыхания изолированных гепатоцитов у птиц с разной массой тел [87].
При расчете литературных данных принято, что в гепатоцитах масса воды составляет 70%.
Похожий процесс происходит в ряду млекопитающих «крыса-кролик-овца-бык»: относительная масса печени уменьшается (≈ Мт-0.18), в ней падает плотность Мх/г (≈ Мт-0.1) и, как следствие, общее количество Мх в печени снижается в 2 раза (≈ Мт-0.3) (Smith, 1956), по: [73].
По мере увеличения Мт теплокровных меняется структура поверхностных мембран в клетках. Эти двуслойные белково-липидные мембраны, содержащие клеточные рецепторы и каналы для обменивающихся веществ, играют очень важную роль в регуляции клеточного метаболизма, в частности окислительного. Оказалось, что с увеличение Мт число ненасыщенных связей в мембранных фосфолипидах снижается, что может приводить к падению скорости транспорта веществ через клеточные мембраны, рис. 18.
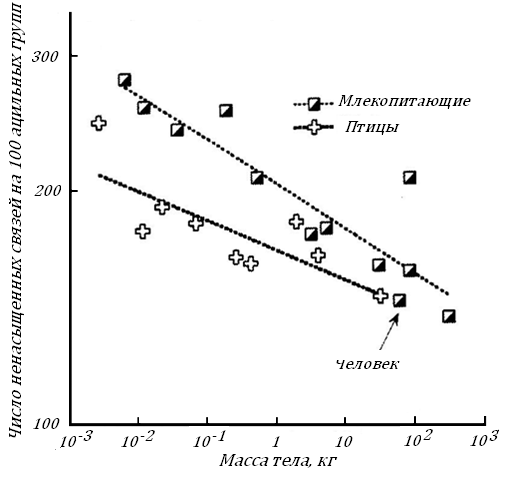
Рис. 18. Число ненасыщенных связей в фосфолипидах клеточных мембран скелетных мышц млекопитающих (А) и птиц (В) с разной массой тела [92].
Таким образом, падение БМ, необходимое по мере увеличения размера животного и уменьшения относительной поверхности его тела, сопровождается снижением скоростей кровотока и окислительных реакций в различных органах. У теплокровных в органах с высокой интенсивностью этих реакций (как правило, они несут вегетативные функции) такое снижение менее выражено, чем в органах с низкой интенсивностью, к которым в условиях БМ относятся опорно-двигательные структуры. Одновременно относительная масса первых органов снижается, а вторых - не меняется или даже растет. При этом альвеолярная поверхность легких, через которую происходит газообмен теплокровного с внешней средой, увеличивается почти пропорционально увеличению Мт.
Задача 4
Найти свой базальный метаболизм:
БМ = 65.8 Мт0.75∙ [1 – 0.001 (30-А) + 0.018 (В – 42.1)]
БМ – ккал/сут; Мт – кг; А - возраст, годы; В – «рост, см / МТ0.33,кг»
Решение задачи 4
Например, у женщины с Мт - 60 кг (600.75 = 21.6), А - 25 лет, рост - 165 см,
В - 165/600.33 = 42.7:
БМ = 65.821.6[1 – 0.0015 + 0.018(42.7 – 42.1)] = 1430∙ккал/сут.
Задача 5. Найти свой идеальный вес по формуле Броке:
Мт, кг = (Рост, см Объем груди, см) / 240
(или Рост –100 – 10%)
Задача 6. У человека идеальная Мт 60 кг, а избыточная масса жировой ткани составляет 20% (вся Мт = 60кг+12кг). Известно, что калорийность жира равна 9.4 ккал/г, а количество воды в жировой ткани 60%. Сколько дней этот человек может жить только за счет своего избыточного жира, если его суточная потребность в энергии будет составлять 1.5 БМ ?
Решение задачи 6. БМ для человека с Мт 60-кг 1509 ккал/сут. Вся потребность в энергии 2264 ккал/сут. Количество жира 12 кг 0.4 = 4.8 кг.
При окислении этого жира выделится 9.4 ккал 4800 г = 45120 ккал
45120 ккал / 2264 ккал/сут = 20 сут
Задача 7. Найти относительную массу органов, % (Мт в кг)
мышь 20 г , человек 60 кг, бык 700 кг
Печень
3.33 Мт–0,13 1.66 0.59 0.43
Почки
0.73 Мт–0,15 1.80 0.54 0.37
Сердце
0.58 Мт–0,02 1.08 0.92 0.88
Решение задачи 7. Относительная масса ,% , (Мт в кг)
мышь 20 г , человек 60 кг, бык 700 кг
Печень 5.53 1.96 1.43
Почки 1.31 0.39 0.27
Сердце 0.63 0.53 0.51
Задача 8. Найти частоты в минуту (Мт в кг)
дыхания 53.5 • Мт–0,26
сердцебиения 241• Мт–0,25
мышь 20 г человек 60 кг бык 700 кг
Мт–0.26 2.77 0.34 0.18
Мт–0.25 2.66 0.36 0.19