

Фотопериодическая реакция и температура
В предыдущих разделах мы уже останавливались на некоторых примерах взаимодействия ФПР и температуры. У большинства насекомых с длиннодневным типом фотопериодической реакции повышение температуры приводит к уменьшению критического фотопериода и устранению диапаузы (рис. 14). На каждые 5° критический фотопериод сдвигается на 1–1,5 ч, что соответствует естественным изменениям фотопериода за 2–4 недели. В результате увеличивается срок, в течение которого возможно развитие насекомого. При температурах, близких к сублетальным, развитие идет практически всегда без диапаузы. Такая зависимая от температуры лабильность ФПР дает возможность некоторым видам проникать из умеренной зоны в субтропическую, где фотопериод всегда непродолжителен. Наоборот, "усиление" ФПР при низкой температуре способствует сохранению вида при необычно раннем наступлении осени.
У видов с короткодневной ФПР, как и следует ожидать, реакция на температуру противоположна и высокая температура способствует наступлению диапаузы.
В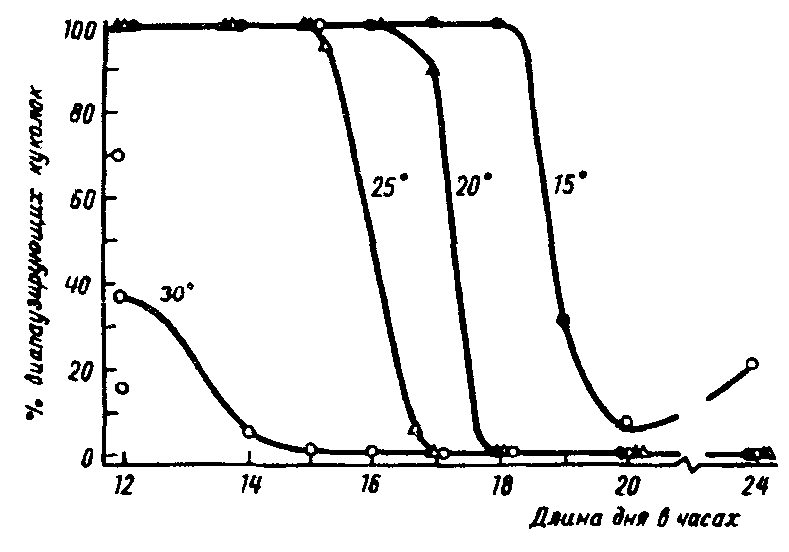 озможны
и другие варианты воздействия температуры
на ФПР. Так, у восточной плодожоркиGrapholitha molestaBusck. и хлопковой совкиHelicoverpa armigera(Hubner) диапауза может
быть вызвана коротким фотопериодом
только в диапазоне температур 20–25°.
Такая связь индукции диапаузы с довольно
высокой температурой скорее всего
объясняется южным происхождением этих
видов. Наоборот, у бабочки–медведицыSpi–losoma menthastriEsp. развитие без
диапаузы, зависящее также от ФПР,
возможно только в пределах температур
20–27°.
озможны
и другие варианты воздействия температуры
на ФПР. Так, у восточной плодожоркиGrapholitha molestaBusck. и хлопковой совкиHelicoverpa armigera(Hubner) диапауза может
быть вызвана коротким фотопериодом
только в диапазоне температур 20–25°.
Такая связь индукции диапаузы с довольно
высокой температурой скорее всего
объясняется южным происхождением этих
видов. Наоборот, у бабочки–медведицыSpi–losoma menthastriEsp. развитие без
диапаузы, зависящее также от ФПР,
возможно только в пределах температур
20–27°.
Рис.14. Зависимость критической длины светового дня у щавелевой совки Acronycta rumicis L. от температуры, при которой происходило развитие гусениц (по А.С.Данилевскому, 1961)
Итак, ФПР обычно имеет место в диапазоне температур, оптимальных для роста и развития. В некоторых случаях, однако, ФПР представляется совсем не связанной с уровнем температуры. С другой стороны, известен ряд видов, у которых наступление диапаузы определяется прежде всего температурой. Подобное влияние температуры четко выражено у таких объектов, как, например, живущие в почве личинки некоторых пластинчатоусых жуков, у находящихся в почве яиц саранчовых. К числу таких насекомых относятся также обитающие в древесине или под корой личинки жуков, некоторые вредители запасов, например гусеницы бабочки Plodia interpunctellaHb., a также ряд тропических видов насекомых.
Как и в случае с фотопериодом, на развитие может влиять не только сам уровень температуры, но и направление ее изменений. Так, повышение температуры уменьшает процент диапаузирующих личинок мух Sarcophaga иCalliphora.Понижение температуры в сочетании с уменьшением длины светового дня резко увеличивает склонность куколок бабочки Heliothis zea к диапаузе.
Суточный ход температуры может также влиять на сезонное развитие. Так, стеблевой кукурузный мотылек Ostrinia nubilalisHb. на 100% диапаузирует при коротком фотопериоде и постоянной температуре, а также при температурном цикле 31° днем и 21° ночью. Если же цикл температуры инвертирован (31° ночью и 21° днем), то доля диапаузирующих особей резко падает, при любом фотопериоде (S.Beck, 1962). Следовательно, индукция диапаузы зависит от уровня температуры в ночное время.
При содержании насекомых в постоянной темноте продолжительность теплого периода в течение суток может играть роль, аналогичную фотопериоду. Здесь кривая индукции диапаузы в зависимости от продолжительности термопериода очень сходна с кривой ФПР.