

Патологическая анатомия Серов
.pdfРис.
свидетельствует как о повышенном синтезе р-РНК, так и о трансмиссии. Такие «гиперфункциональные ядрышки» встречаются в молодых плазматических клетках, активных фибробластах, гепатоцитах, во многих опухолевых клетках. Те же гипергранулированные ядрышки со слабовыраженной базофилией цитоплазмы могут отражать нарушение трансмиссии (транспортировки гранул) при продолжающемся синтезе р-РНК. Они обнаруживаются в опухолевых клетках, отличающихся большим ядром и незначительной цитоплазматической базофилией.
Разрыхление (диссоциация) ядрышек, отражающее их гипогрануляцию, может быть следствием «извержения» р-РНК в цитоплазму или торможения ядрышковои транскрипции. Дезорганизация (сегрегация) ядрышекотражает, как правило, полное и быстрое прекращение ядрышковой транскрипции: ядро уменьшается в размерах, наблюдается выраженная конденсация ядрышкового хроматина, происходит разделение гранул и протеиновых нитей. Эти изменения встречаются при энергетическом дефиците клетки.
Ядерные включения
Ядерные включения делят на три группы: ядерные цитоплазматические, истинные ядерные и ядерные вирусобусловленные.
Ядерными цитоплазматическими включениями называют отграниченные оболочкой части цитоплазмы в ядре. Они могут содержать все составные части клетки (органеллы, пигмент, гликоген, капли жира и т.д.). Их появление в большинстве случаев связано с нарушением митотического деления.
21
Истинными ядерными включениями считают те, которые расположены внутри ядра (кариоплазмы) и соответствуют веществам, встречающимся
в цитоплазме - белок, гликоген (рис. 6, а), липиды и т.д. В большинстве случаев эти вещества проникают из цитоплазмы в ядро через неповрежденные или поврежденные поры ядерной оболочки или через разрушенную ядерную оболочку. Возможно также проникновение этих веществ в ядро при митозе. Таковы, например, включения гликогена в ядрах печени при сахарном диабете («ядерный гликоген», «дырчатые, пустые, ядра»).
22
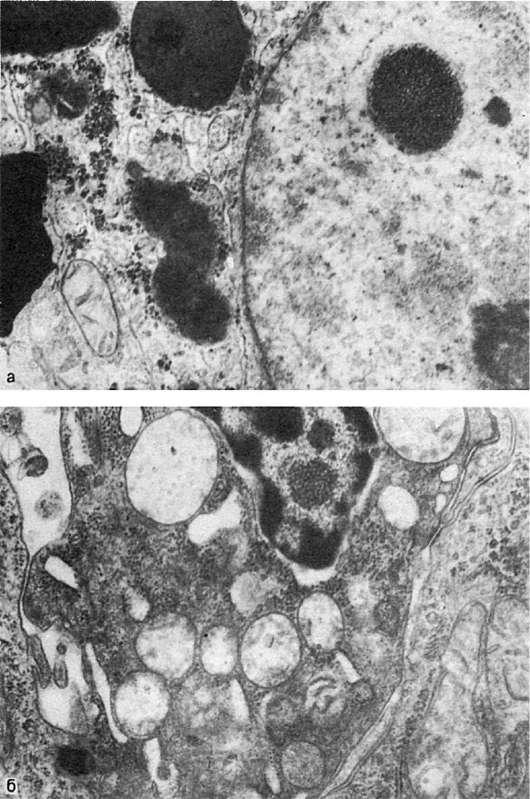
Рис.
а - включения гликогена в ядре гепатоцита. х22 500; б - включения вируса в ядре опухолевой клетки
Вирусобусловленные ядерные включения (так называемые тельца ядерных включений) неоднозначны. Во-первых, это ядерные включения в кариоплазме кристаллической решетки
23
вируса (рис. 6, б), во-вторых, включения белковых частиц, возникающих при внутриядерном размножении вируса; в-третьих, ядерные включения как проявление реакции на поражение вирусом цитоплазмы («реактивные включения»).
Ядерная оболочка
Ядерная оболочка выполняет ряд функций, нарушения которых могут служить основой для развития патологии клетки.
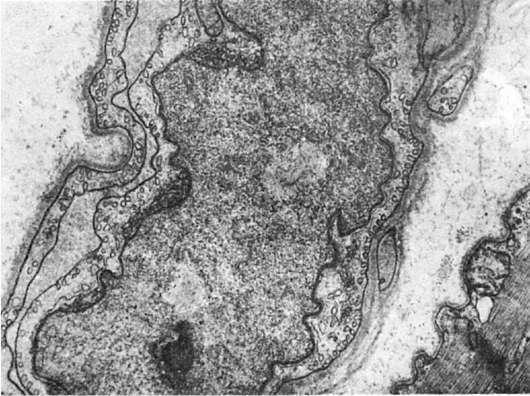
Ороли ядерной оболочки в поддержании формы и размера ядра свидетельствует образование внутриядерных трубчатых систем, отходящих от внутренней ядерной мембраны, включений в перинуклеарной зоне - гипертрофия миокарда, легочный фиброз, системный васкулит, саркоидоз, опухоли печени, дерматомиозит (рис. 7).
Оядерной оболочке как месте прикрепления ДНК для облегчения репликации и транскрипциисвидетельствует тот факт, что в ядерной оболочке имеются структуры, модулированные хроматином и в свою очередь ответственные за ориентацию и структуру хроматина. Показано, что функциональная активность ДНК связана с ее распределением при делении клетки и со степенью конденсации в интерфазе, причем повреждение оболочки может вызывать изменения таких участков распределения и быть причиной патологических изменений клетки.
Рис.
7. Микротубулярные включения в перинуклеарной зоне эндотелиоцита при дерматомиозите.
х15 500
В пользу функции ядерной оболочки как физического барьера и модулятора нуклеоцитоплазматического обмена говорит установленная корреляция между
24
изменениями структуры ядерной оболочки, модулем ее пор и выходом РНК в цитоплазму. Контроль ядерной оболочкой транспорта РНК в цитоплазму может оказывать существенное влияние на гомеостаз клетки при патологических состояниях. Участие ядерной оболочки всинтезе мембран не имеет достоверных доказательств, хотя и считают, что эта роль возможна, так как мембраны ядерной оболочки непосредственно переходят в эндоплазматическую сеть цитоплазмы. О возможном влиянии ферментов ядерной оболочки на функцию ядра свидетельствует наличие в ядерной оболочке различных ферментов детоксикации, а также веществ, обеспечивающих «гормональное управление» (аденилатциклаза, рецепторы инсулина и др.).
Патология митоза
В жизненном цикле клетки митоз занимает особое место. С его помощью осуществляется репродукция клеток, а значит, и передача их наследственных свойств. Подготовка клеток к митозу складывается из ряда последовательных процессов: репродукции ДНК, удвоения массы клетки, синтеза белковых компонентов хромосом и митотического аппарата, удвоения клеточного центра, накопления энергии для цитотомии. В процессе митотического деления, как известно, различают 4 основные фазы: профазу, метафазу, анафазу и телофазу.
При патологии митоза может страдать любая из этих фаз. Руководствуясь этим, создана классификация патологии митоза (Алов И.А., 1972), согласно которой выделяются следующие типы патологии митоза.
I. Повреждение хромосом: 1) задержка клеток в профазе; 2) нарушение спирализации и деспирализации хромосом; 3) фрагментация хромосом; 4) образование мостов между хромосомами в анафазе; 5) раннее разъединение сестринских хроматид; 6) повреждение кинетохора.
II. Повреждение митотического аппарата: 1) задержка развития митоза в метафазе; 2) рассредоточение хромосом в метафазе; 3) трехгрупповая метафаза; 4) полая метафаза; 5) многополюсные митозы; 6) асимметричные митозы; 7) моноцентрические митозы; 8) К- митозы.
III. Нарушение цитотомии: 1) преждевременная цитотомия; 2) задержка цитотомии; 3) отсутствие цитотомии.
Патологию митоза могут вызвать различные воздействия на клетку: ультрафиолетовое и ионизирующее излучение, высокая температура, химические вещества, в том числе канцерогены и митотические яды и др. Велико количество патологических митозов при малигнизации тканей (рис. 8).
25
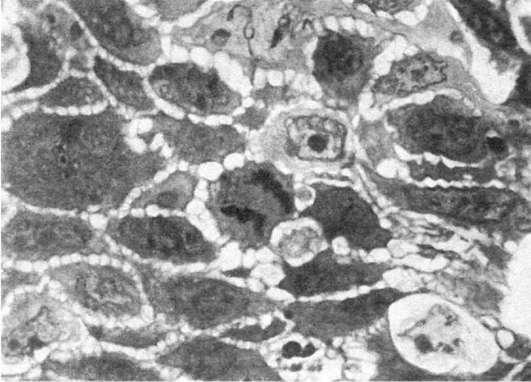
Рис.
8. Патология митоза. Полутонкий срез ткани опухоли. х1000
Хромосомные аберрации и хромосомные болезни
Хромосомные аберрации. Под хромосомными аберрациями понимают изменения структуры хромосом, вызванные их разрывами, с последующим перераспределением, утратой или удвоением генетического материала. Они отражают различные виды аномалий хромосом. У человека среди наиболее часто встречающихся хромосомных аберраций, проявляющихся развитием глубокой патологии, выделяют аномалии, касающиеся числа и структуры хромосом. Нарушения числа хромосом могут быть выражены отсутствием одной из пары гомологичных хромосом (моносомия) или появлением добавочной, третьей, хромосомы (трисомия).Общее количество хромосом в кариотипе в этих случаях отличается
от модального числа и равняется 45 или 47. Полиплоидия и анеуплоидия имеют меньшее значение для развития хромосомных синдромов. К нарушениям структуры хромосом при общем нормальном их числе в кариотипе относят различные типы их «поломки»:
транслокацию (обмен сегментами между двумя негомологичными хромосомами), делецию (выпадение части хромосомы), фрагментацию, кольцевые хромосомы и т.д.
Хромосомные аберрации, нарушая баланс наследственных факторов, являются причиной многообразных отклонений в строении и жизнедеятельности организма, проявляющихся в так называемых хромосомных болезнях.
Хромосомные болезни. Их делят на связанные с аномалиями соматических хромосом (аутосом) и с аномалиями половых хромосом (телец Барра). При этом учитывают характер хромосомной аномалии - нарушение числа отдельных хромосом, числа хромосомного набора или структуры
26
хромосом. Эти критерии позволяют выделять полные или мозаичные клинические формы хромосомных болезней.
Хромосомные болезни, обусловленные нарушениями числа отдельных хромосом (трисомиями и моносомиями), могут касаться как аутосом, так и половых хромосом.
Моносомии аутосом (любые хромосомы, кроме Х- и Y-хромосом) несовместимы с жизнью. Трисомии аутосом достаточно распространены в патологии человека. Наиболее часто они представлены синдромами Патау (13-я пара хромосом) и Эдвардса (18-я пара), а также болезнью Дауна (21-я пара). Хромосомные синдромы при трисомиях других пар аутосом встречаются значительно реже. Моносомия половой Х-хромосомы (генотип ХО) лежит в основе синдрома Шерешевского-Тернера, трисомия половых хромосом (генотип XXY) - в основе синдрома Клейнфелтера. Нарушения числа хромосом в виде тетраили триплоидии могут быть представлены как полными, так и мозаичными формами хромосомных болезней.
Нарушения структуры хромосом дают самую большую группу хромосомных синдромов (более 700 типов), которые, однако, могут быть связаны не только с хромосомными аномалиями, но и с другими этиологическими факторами.
Для всех форм хромосомных болезней характерна множественность проявлений в виде врожденных пороков развития, причем их формирование начинается на стадии гистогенеза и продолжается в органогенезе, что объясняет сходство клинических проявлений при различных формах хромосомных болезней.
Патология цитоплазмы
Изменения мембран и патология клетки
Клеточные мембраны, как известно, состоят из бислоя фосфолипидов, по обе стороны которого располагаются разнообразные мембранные белки. На внешней поверхности мембраны белковые молекулы несут полисахаридные компоненты (гликокаликс), которые содержат многочисленные поверхностные клеточные антигены. Они играют важную роль в клеточном узнавании, формировании клеточных стыков.
Изменения клеточных мембран. Среди них различают следующие (Авцын А.П., Шахламов В.А., 1979): чрезмерное везикулообразование («минусмембрана» - рис. 9); увеличение поверхности плазмолеммы клеток мембранами микропиноцитозных пузырьков («плюсмембрана»); усиленный микроклазматоз и клазматоз («минус-мембрана» - см. рис. 9); образование цитоплазматических отростков из плазмолеммы клетки; образование пузырей на поверхности клетки; утолщение слоев мембраны; образование микропор; образование миелиноподобных структур из плазмолеммы и мембран органелл; слияние разнородных клеточных мембран; локальные разрушения мембран - «бреши» в плазмолемме; «штопка» локально разрушенной плазмолеммы мембранами микропиноцитозных везикул.
27
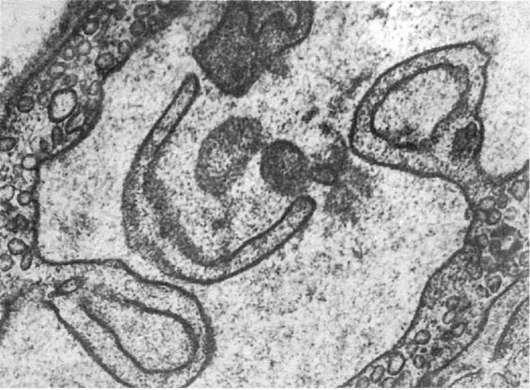
Рис.
9. Изменения мембран эндотелиоцитов. Усиленное везикулообразование и клазматоз. х25
500
К патологии мембран клетки могут вести нарушения мембранного транспорта, изменения проницаемости мембран, изменения коммуникации клеток и их «узнавания», изменения подвижности мембран и формы клеток, нарушения синтеза и обмена мембран.
Нарушения мембранного транспорта. Процесс мембранного транспорта предполагает перенос ионов и других субстратов против градиента концентрации. Транспорт может быть активным, тогда он требует АТФ и «подвижности» транспортных белков в мембране, или пассивным посредством различных диффузионных и обменных процессов. Активный транспорт - это также функция эпителиальных барьеров. Нарушения мембранного транспорта, ведущие к патологии клетки, хорошо прослежены при ишемии, которая приводит к первичным изменениям митохондрий. В митохондриях резко падает эффективность окислительного фосфорилирования, они набухают, вначале увеличивается проницаемость их внутренней мембраны, в дальнейшем повреждение становится тотальным и необратимым
(рис. 10).
Ишемическое повреждение митохондрий приводит к полому натрий-калиевого АТФ-насоса, постепенному накапливанию в клетке натрия и потере ею калия. Нарушение натрий-калиевого обмена ведет к вытеснению кальция из митохондрий. В результате в цитоплазме повышается уровень ионизированного кальция и увеличивается связывание его с кальмодулином. С повышением содержания кальцийкальмодулиновых комплексов связан ряд изменений клетки: рас-
28
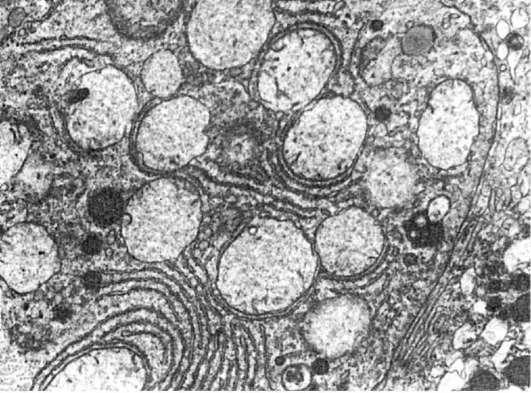
Рис.
10. Вакуолизация митохондрий в одном гепатоците (слева) и конденсация их в другом
(справа). х16 000
хождение клеточных стыков, поглощение кальция митохондриями, изменение микротрубочек и микрофиламентов, активация фосфолипаз. Эндоплазматическая сеть накапливает воду и ионы, следствием чего является расширение ее канальцев и цистерн, развитие гидропической дистрофии. Усиление гликолиза сопровождается истощением гликогена, накоплением лактата и снижением клеточного рН. С этими изменениями связано нарушение структуры хроматина и уменьшение синтеза РНК. Необратимые ишемические повреждения клетки связаны с гидролизом мембран, особенно мембранных липидов, под действием фосфолипаз. Возникают и нарушения лизосомальных мембран с высвобождением гидролаз.
Изменения проницаемости мембран. Контроль мембранной проницаемости предполагает поддержание структуры как фосфолипидного бислоя мембраны с необходимым обменом и ресинтезом, так и соответствующих белковых каналов. Важная роль в осуществлении этого контроля принадлежит гликокаликсу и взаимодействию мембранных белков с цитоскелетом, а также гормонам, взаимодействующим с мембранными рецепторами. Изменения проницаемости могут быть тяжелыми (необратимыми) или поверхностными. Наиболее изученной моделью изменения мембранной проницаемости является повреждение тяжелыми металлами (ртуть, уран). Тяжелые металлы, взаимодействуя с сульфгидрильными группами мембранных белков, изменяют их конформацию и резко увеличивают проницаемость мембраны для натрия,
калия, хлора, кальция и магния, что приводит к быстрому набуханию клеток, распаду их цитоскелета. Подобные изменения мембран отмечаются при повреждении их комплементом
29
(«болезни гиперчувствительности»). В мембранах образуются бреши, что снижает их сопротивление и резко увеличивает проницаемость.
Изменения коммуникации клеток и их «узнавания». Коммуникабельность клеток и опознавание «своих» и «чужих» - необходимое свойство клеточного кооперирования. Клеточное «общение» и «узнавание» подразумевают прежде всего различия во внешних поверхностях плазматической мембраны и мембран внутриклеточных органелл. Особый интерес в этом отношении представляет гликокаликс мембраны с поверхностными антигенами - маркерами определенного типа клеток.
Изменения клеточного «общения» и «узнавания» встречаются при тех патологических процессах (воспаление, регенерация, опухолевый рост), при которых поверхностные антигены могут изменяться, причем различия могут касаться как типа антигена, так и его «доступности» со стороны внеклеточного пространства. Показано, что при исчезновении характерных для данного типа клеток антигенов могут появляться «эмбриональные» и аномальные (например, карциноэмбриональный) антигены; изменения гликолипидов мембраны делают ее более доступной воздействию антител.
Коммуникабельность клеток определяется также состоянием клеточных стыков, которые могут повреждаться при различных патологических процессах и болезнях. В раковых клетках, например, найдена корреляция между изменениями клеточных стыков и нарушением межклеточных связей; в опухолях обнаружены аномальные клеточные соединения.
Изменения подвижности мембран и формы клеток. Различают два типа изменений,
связанных с нарушением подвижности мембран: выпячивание мембраны наружу - экзотропия и внутрь цитоплазмы - эзотропия. При экзотропии мембрана, выпячивающаяся во внеклеточное пространство, образует окруженную мембраной цитоплазматическую структуру. При эзотропии появляется окруженная мембраной полость. Изменения формы клеток связаны не только с экзо- и эзотропией, но и с упрощением клеточной поверхности (потеря малых отростков подоцитов при нефротическом синдроме).
Нарушения синтеза и обмена мембран. Возможно усиление синтеза мембран (при воздействии ряда химических веществ на клетку) или его ослабление (снижение синтеза мембран щеточной каемки энтероцитов при угнетении мембранных ферментов). В равной мере возможно усиление обмена мембран (при стимуляции аутофагоцитоза) или его ослабление (при лизосомных болезнях).
Эндоплазматическая сеть
Однозначные изменения гранулярной и агранулярной эндоплазматической сети могут отражать нарушения различных функций цитоплазмы и клетки.
Изменения гранулярной эндоплазматической сети и рибосом
Функции гранулярной эндоплазматической сети и рибосом сопряжены достаточно жестко, поэтому морфологические проявления их нарушений касаются, как правило, обеих органелл.
30