

2.2. Физиологические свойства скелетных мышц. Фазовые изменения возбудимости нервной и мышечной ткани. Методы измерения возбудимости Физиологические свойства мышц
• Возбудимость - способность приходить в состояние возбуждения при действии раздражителей.
• Проводимость - способность проводить возбуждение.
• Сократимость - способность мышцы изменять свою длину или напряжение в ответ на действие раздражителя.
• Лабильность – по Н.Е.Введенскому, наибольшее число потенциалов действия, которое возбудимая ткань способна воспроизвести в единицу времени (1 сек.) под влиянием частых приложений к ней раздражений (лабильность мышечного волокна равна 20-30 импульсов в секунду, нервного около 1000).
Автоматия – способность генерировать импульсы без внешнего раздражения (свойство характерно для сердечной мышцы и для гладких мышц).
Скелетные (поперечнополосатые) мышцы в организме играют роль своеобразных «машин», преобразующих химическую энергию непосредственно в механическую и тепловую. Сокращение мышц возникает в ответ на электрические импульсы, приходящие к ним от альфа-мотонейронов – нервных клеток, лежащих в передних рогах спинного мозга.
Мышцы и иннервирующие их мотонейроны составляют нервно-мышечный аппарат человека.
Организм человека обладает выраженной способностью адаптироваться к постоянно меняющимся условиям внешней среды. В основе приспособительных реакций организма лежит универсальное свойство живой ткани - раздражимость - способность отвечать на действие раздражающих факторов изменением структурных и функциональных свойств. Раздражимостью обладают все ткани животных и растительных организмов. В процессе эволюции происходила постепенная дифференциация тканей, осуществляющих приспособительную деятельность организма. Раздражимость этих тканей достигла наивысшего развития и трансформировалась в новое свойство - возбудимость. Под этим термином понимают способность ткани отвечать на раздражение специализированной реакцией - возбуждением. Возбуждение - это сложный биологический процесс, который характеризуется специфическим изменением процессов обмена веществ, теплообразования, временной деполяризацией мембраны клеток и проявляющийся специализированной реакцией ткани (сокращение мышцы, отделение секрета железой и т. д.).
Возбудимостью обладают нервная, мышечная и секреторная ткани, их объединяют в понятие "возбудимые ткани".
Возбудимость различных тканей неодинакова. Мерой возбудимости является порог раздражения - минимальная сила раздражителя, которая способна вызвать возбуждение. Менее сильные раздражители называются подпороговыми, а более сильные - сверхпороговыми. Раздражителем живой клетки может быть любое изменение внешней или внутренней среды, если оно достаточно велико, возникло достаточно быстро и продолжается достаточно долго.
Природа возбуждения
Первые попытки последовательной разработки учения о "животном электричестве" связаны с именем Л. Гальвани. Э. Дюбуа-Реймон впервые показал, что наружная поверхность мышцы заряжена положительно по отношению к ее внутреннему содержимому. Следовательно, в состоянии покоя между наружной и внутренней поверхностями мембраны клетки существует разность потенциалов, которая затем была названа мембранным потенциалом покоя или мембранным потенциалом. Его величина у разных клеток колеблется от 60 до 90 мВ.
А. Ходжкин, А. Хаксли и Б. Катц в 50-х годах 20 века объяснили причины возникновения мембранного потенциала покоя, для чего они существенно переработали ранее существовавшие представления и создали мембранно-ионную теорию. Согласно их взглядам мембранный потенциал покоя (МПП) обусловлен неодинаковой концентрацией ионов натрия, калия, кальция, хлора внутри клетки и во внеклеточной жидкости, а также неодинаковой проницаемостью для этих ионов поверхностной мембраны клетки (рис. 2.4). Цитоплазма нервных и мышечных клеток содержит в 30-50 раз больше ионов калия, в 8-10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Следовательно, в состоянии покоя существует асимметрия концентрации ионов внутри клетки и в окружающей ее среде.

Рис. 2.4. Регистрация потенциала покоя
В мембране имеются ионные каналы, образованные макромолекулами белка, пронизывающих липидный слой. Каналы мембраны делятся на неспецифические (каналы утечки) и специфические (селективные, обладающие способностью пропускать только определенные ионы). Неспецифические каналы пропускают различные ионы и открыты постоянно. Специфические каналы открываются и закрываются в ответ на изменения МПП. Эти каналы называются потенциалозависимыми.
В состоянии физиологического покоя мембрана нервных волокон в 25 раз лучше проницаема для K+, чем для Na+.
Выход положительно заряженных ионов калия приводит к появлению положительного заряда на наружной поверхности мембраны. Органические анионы - крупномолекулярные соединения, которые несут отрицательный заряд, и для которых мембрана клетки непроницаема, придают в этих условиях внутренней поверхности мембраны отрицательный заряд (рис.2.5).
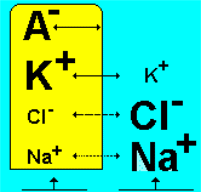
Рис.2.5. Концентрация основных ионов внутри и вне клетки.
В состоянии покоя существуют небольшие перемещения K+и Na+ через мембрану по их концентрационному градиенту (табл.2.2), K+больше, чем Na+.
Табл. 2.2.
Концентрационные градиенты основных ионов клетки в покое
|
Концентрация ионов, мМ/л |
|
|
Внеклеточная среда |
Цитоплазма клетки |
Na+ |
144 |
10 |
K+ |
4 |
140-160 |
Cl− |
114 |
2 - 25 |
Такие процессы в конечном итоге должны были бы привести к уравновешиванию концентраций ионов между клеткой и окружающей средой. Однако в живом организме этого не происходит благодаря постоянной работе калий-натриевого насоса. Этот молекулярный механизм обеспечивает выведение из цитоплазмы клетки Na+ и перемещение в цитоплазму K+. Ионный насос перемещает ионы против их концентрационного градиента, поэтому для его работы необходима затрата энергии.
Регистрация электрических потенциалов в нервном и мышечном волокне или в нервной клетке показала, что при возбуждении происходит изменение МПП, возникает потенциал действия. Под влиянием раздражителя пороговой или сверхпороговой величины проницаемость мембраны клетки для ионов натрия возрастает. Ионы Na+ устремляются внутрь клетки, что приводит к уменьшению величины мембранного потенциала покоя - деполяризация мембраны (рис. 2.6.).
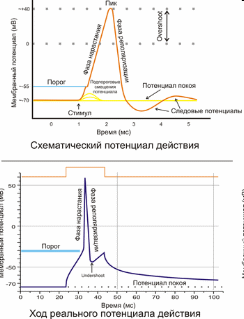
Рис. 2.6. Потенциал действия (схема и реальный процесс - http://dic.academic.ru/dic.nsf/ruwiki/227096).
Вначале деполяризация развивается медленно. При уменьшении МПП до критического уровня деполяризации проницаемость мембраны для Na+ увеличивается в 500 раз и превышает проницаемость для K+ в 20 раз. В результате проникновения Na+ в цитоплазму и их взаимодействия с анионами разность потенциалов на мембране исчезает, а затем происходит перезарядка клеточной мембраны (инверсия заряда) - внутренняя поверхность мембраны заряжается положительно по отношению к ее наружной. Этот потенциал превышения достигает величины 30-50 мВ, после чего закрываются быстрые натриевые каналы - происходит инактивация натриевой проницаемости и открываются калиевые каналы, в работу активно включается калий-натриевый насос. Начинается процесс восстановления исходного уровня мембранного потенциала покоя - реполяризация мембраны.
При внутриклеточном отведении регистрируются следующие состояния мембраны (фазы потенциала действия):
- местное возбуждение, локальный ответ (начальная деполяризация мембраны);
- деполяризация мембраны (восходящая часть потенциала действия, включая инверсию);
- реполяризация мембраны (нисходящая часть потенциала действия);
- следовая деполяризация (соответствует отрицательному следовому потенциалу);
- следовая гиперполяризация (соответствует положительному следовому потенциалу).
Изменение возбудимости при возбуждении.
При развитии потенциала действия происходит изменение возбудимости ткани, причем, это изменение протекает по фазам (рис. 2.7).
Состоянию исходной поляризации мембраны, которую отражает мембранный потенциал покоя, соответствует исходное состояние ее возбудимости и, следовательно, клетки - это нормальный уровень возбудимости. В период локального ответа возбудимость ткани повышена, эта фаза возбудимости получила название первичной экзальтации. Во время развития локального ответа мембранный потенциал покоя приближается к критическому уровню деполяризации и для достижения последнего достаточна сила раздражителя меньшая, чем пороговая (подпороговая).
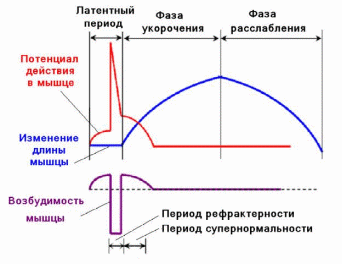
Рис. 2.7. Соотношение одиночного цикла возбуждения (А) и фаз возбудимости (Б), (по www.satisform.ru/cat/smi5344344).
А: а - мембранный потенциал покоя; б - предспайк, локальный ответ, ВПСП; в - спайк, потенциал действия, деполяризация и инверсия; г - потенциал действия, реполяризация; д - отрицательный следовой потенциал, следовая деполяризация; е - положительный следовой потенциал, следовая гиперполяризация. Б: а - исходный уровень возбудимости; б - фаза первичной экзальтации, повышенная возбудимость; в - фаза абсолютной рефрактерности; г - фаза относительной рефрактерности; д - фаза вторичной экзальтации; е - фаза вторичной рефрактерности
Начало развития пикового потенциала сопровождается лавинообразным поступлением Na+ внутрь клетки, заряд мембраны изменяется на противоположный (инверсия заряда). В этот момент ее способность отвечать возбуждением утрачивается даже на сверхпороговые раздражители до конца перезарядки мембраны. Эта фаза возбудимости называется абсолютной рефрактерностью (абсолютной невозбудимостью). Возникновение этой фазы связано с тем, что сначала натриевые каналы полностью открываются, а затем происходит их инактивация.
После завершения фазы инверсии начинается восстановление возбудимости мембраны до исходного уровня. Развивается фаза относительной рефрактерности. Она продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Так как в этот период мембранный потенциал покоя еще не восстановлен, то возбудимость ткани понижена, и новое возбуждение может возникнуть только при действии сверхпорогового раздражителя. Снижение возбудимости в фазу относительной рефрактерности связано с частичной инактивацией натриевых каналов и активацией калиевых.
Периоду отрицательного следового потенциала соответствует повышенный уровень возбудимости - фаза вторичной экзальтации. Так как мембранный потенциал в эту фазу ближе к критическому уровню деполяризации, но сравнению с состоянием покоя (исходной поляризацией), то порог раздражения снижен, т. е. возбудимость повышена. В эту фазу новое возбуждение может возникнуть при действии раздражителей подпороговой силы. Натриевые каналы в эту фазу инактивированы не полностью. В период развития положительного следового потенциала возбудимость ткани понижена - фаза вторичной рефрактерности. В эту фазу мембранный потенциал увеличивается (состояние гиперполяризации мембраны), удаляясь от критического уровня деполяризации, порог раздражения повышается, таким образом, новое возбуждение может возникнуть только при действии раздражителей сверхпороговой величины. Гиперполяризация мембраны развивается вследствие трех причин: во-первых, продолжающимся выходом ионов K+; во-вторых, открытием, возможно, каналов для анионов Cl− и поступление этих ионов в цитоплазму клетки; в-третьих, усиленной работой калий-натриевого насоса.
При раздражении с помощью постоянного тока отмечается следующая закономерность: эффект (возбуждение) зависит не только от величины тока, но и от времени его действия - закон силы-длительности. Чем больше ток, тем меньше времени он должен действовать для возникновения возбуждения. Зависимость силы-длительности имеет гиперболический характер (рис.2.8).

Рис. 2.8. Графическое выражение закона силы-длительности (кривая Гоорвега-Вейса-Лапика). 0-1 - реобаза, 0-3 - хронаксия, 0-4 - полезное время, 0-2 две реобазы; по оси абсцисс - длительность раздражения, по оси ординат - сила раздражения
Следствием этой закономерности является то, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и чем короче импульсы тока, тем меньшую раздражающую способность они имеют. Причиной этого электрическая емкость мембраны. Токи малой длительности не успевают разрядить эту емкость (уменьшить разность потенциалов) до критического уровня деполяризации. Минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии, называется реобазой. Время, в течение которого действует ток, равный реобазе, и вызывает возбуждение, называется полезным временем.
В связи с тем, что определение этого времени затруднено, было введено понятие хронаксия - минимальное время, в течение которого ток, равный двум реобазам, должен действовать на ткань, чтобы вызвать ответную реакцию. Определение хронаксии - хронаксиметрия - находит применение в клинике. Электрический ток, приложенный к мышце, проходит через как мышечные, так и нервные волокна, и их окончания, находящиеся в этой мышце. Так как хронаксия нервных волокон значительно меньше хронаксии мышечных волокон, то при исследовании хронаксии мышцы практически получают хронаксию нервных волокон.
Проксимальные мышцы имеют меньшую хронаксию, чем дистальные. Мышца и иннервирующий ее нерв имеют одинаковую хронаксию (изохронизм). Мышцы — синергисты имеют также одинаковую хронаксию. На верхних конечностях хронаксия мышц-сгибателей в два раза меньше хронаксии разгибателей, на нижних конечностях отмечается обратное соотношение. У спортсменов резко снижается хронаксия мышц и может увеличиваться разница хронаксии (анизохронаксия) сгибателей и разгибателей при перетренировке (переутомлении), развитии патологических состояний в мышцах и др.
Величины хронаксии и реобазы не всегда дают представление об истинном состоянии возбудимости нервно-мышечного аппарата. Показателем возбудимости, учитывающим состояние мышцы, является лабильность. Под лабильностью или функциональной подвижностью Н.Е. Введенский понимал большую или меньшую скорость тех элементарных реакций, которыми сопровождается физиологическая деятельность данного аппарата, в частности мышечного. Мерой лабильности по Введенскому является наибольшее число потенциалов действия, которое возбудимый субстрат способен воспроизвести в 1 сек под влиянием раздражителя без трансформации (уменьшения или повышения) ритма раздражения.
Лабильность определяется длительностью пика потенциала действия, т. е. продолжительностью фазы абсолютной рефрактерности. Так как длительность абсолютной рефрактерности у потенциала действия нервного волокна самая короткая, то лабильность его самая высокая. Нервное волокно способно воспроизвести до 1000 импульсов в секунду.