

Продолжительность изучения темы_______________ часов
Из них на занятие ___________ часов; самостоятельная работа_________ часов.
Место проведения учебная комната
Цель занятия знать механизмы формирования мембранных потенциалов в возбудимых тканях; уметь оценить изменения мембранной ионной проницаемости и возбудимости тканей под влиянием различных факторов.
Задачи:
-
знать историю изучения биоэлектрических явлений в живых тканях;
-
знать особенности строения возбудимости мембраны и её физиологические свойства (избирательная ионная проницаемость, ионные каналы, пассивный и активный ионный транспорт);
-
знать механизмы формирования мембранного потенциала покоя;
-
знать механизмы формирования электротонического и местного потенциалов;
-
знать механизмы формирования мембранного потенциала действия;
-
уметь оценить изменения поляризации мембраны при действии раздражителей подпороговой, пороговой сверхпороговой величины;
-
уметь оценить изменения возбудимости ткани во время одиночного цикла возбуждения;
-
уметь выявить наличие токов «покоя» и «действия» в живых тканях.
Методические рекомендации по самоподготовке.
Биологический смысл раздражимости. Возбуждение и его физиологические основы
Неотъемлемым свойством организмов и всех живых систем является раздражимость - способность воспринимать внешние или внутренние раздражители (воздействия) и адекватно на них реагировать. Раздражителем живой клетки может быть любое изменение внешней или внутренней среды, если оно достаточно велико, возникло достаточно быстро и продолжается достаточно долго. Энергией для реакции клетки служит не энергия раздражителя, а энергия, образующаяся в результате метаболизма в самой биологической системе, а сила и форма реакции клетки не определяется силой и формой внешнего воздействия.
Многообразие и сложность ответных реакций многоклеточных животных связана с тем, что в процессе эволюции происходила постепенная дифференциация тканей, осуществляющих приспособительную деятельность организма. Раздражимость этих тканей достигла наивысшего развития и трансформировалась в новое свойство – возбудимость, которая является частным случаем наиболее общего свойства всех клеток — раздражимости. Под возбудимостью понимают способность ткани отвечать на раздражение специализированной реакцией - возбуждением.
Возбуждение - это сложный биологический процесс, который характеризуется специфическим временным изменением заряда мембраны клеток способным распространяться по их поверхности и проявляющийся специализированной реакцией ткани (сокращение секреция и т. д.). Возбуждение – активный процесс, т.е. он может продолжаться и после прекращения действия раздражителя. Ткани способные к возбуждению – объединяют в понятие «возбудимые ткани», их три:
-
нервные клетки (возбуждение проявляется генерацией электрического импульса);
-
мышечные клетки (возбуждение проявляется сокращением);
-
секреторные клетки (возбуждение проявляется выбросом в межклеточное пространство биологически активных веществ);
Клетки возбудимых тканей могут находиться в двух дискретных состояниях: состоянии покоя (готовность к реагированию на внешнее воздействие, совершение внутренней работы) и состоянии возбуждения (активное выполнение специфических функций, совершение внешней работы).
В состоянии покоя мембрана возбудимой клетки поляризована, т.е. имеется постоянная разность потенциалов между внутренней и наружной поверхностью клеточной мембраны, которую называют мембранный потенциал (МП). Значение МП клетки в состоянии покоя называют потенциалом покоя (ПП), его можно измерять, разместив один электрод внутри, а другой снаружи клетки (рис.1 А), В состоянии покоя внутренняя сторона мембраны заряжена отрицательно относительно наружной.
В
состоянии возбуждения происходит
активное изменение мембранного
потенциала. Уменьшение МП относительно
его нормального уровня (ПП) называют
деполяризацией, а
увеличение - гиперполяризацией.
Под реполяризацией
понимают восстановление исходного
уровня МП после его изменения (см. рис.1
Б ).
).
Рис. 1. Схема регистрации мембранного потенциала клетки (А);
мембранный потенциал клетки в состоянии покоя и его возможные изменения (Б).
1 – деполяризация, 2 – гипер-поляризация, 3 – реполяриза-ция.
Наблюдение биоэлектрических явлений. Мембранная теория возбуждения.
В конце XVIII в. (1786) профессор анатомии Болонского университета Луиджи Гальвани провел ряд опытов, положивших начало целенаправленным исследованиям биоэлектрических явлений. Он обратил внимание на сокращение мышц препарата задних лапок лягушки, подвешенного на медном крючке, при прикосновении лапок к железным перилам балкона. На основании этих наблюдений Л. Гальвани пришел к выводу, что сокращение мышц лапок вызвано воздействием на них электричества, источником которого выступают «животные ткани» — мышцы и нервы. Этот опыт в настоящее время известен как первый опыт Гальвани. Физик и физиолог А. Вольта — оспорил это заключение, повторив опыт Гальвани, он пришел к заключению, что они не доказывают наличия "животного электричества". По его мнению, причиной сокращения мышц был электрический ток, возникающий в области контакта двух разнородных металлов: меди и железа (гальваническая пара) — с тканями лягушки.
В ответ на эти возражения Гальвани усовершенствовал опыт, исключив из него металлы, нерв нервно-мышечного препарата набрасывался на специально поврежденную мышцу стеклянным крючком так, чтобы он касался поврежденного и неповрежденного ее участков. В этом случае мышца также сокращалась. Этот опыт известен как второй опыт Гальвани, или опыт без металлов. В этом опыте были получены абсолютные доказательства существования «животного электричества». В 1797 открытие Л. Гальвани подтвердил немецкий натуралист А. Гумбольдт.
Разность электрических потенциалов между поврежденной и неповрежденной частями мышцы обнаружил в 1837 итальянский физик и физиолог Карло Маттеуччи.
Маттеучи произвел также опыт, известный под названием опыта вторичного сокращения: при накладывании на сокращающуюся мышцу нерва второго нервно-мышечного препарата его мышца тоже начинает сокращаться. Результат опыта Маттеучи объясняется тем, что возникающий в мышце при ее возбуждении потенциал действия оказывается достаточно сильным, чтобы вызвать возбуждение другого нерва и мышцы.
В 1848 г. германский физиолог Э. Дюбуа-Реймон, используя более совершенные средства экспериментальной техники, подтвердил существование разности потенциалов между поврежденным и неповрежденным участками невозбужденной мышцы или нерва, а также то, что «ток повреждения» уменьшается при возбуждении. Это «отрицательное колебание» в последствии было названо «потенциалом действия». Работы Э. Дюбуа-Реймона положили начало современной электрофизиологии.
Было разработано несколько теорий возникновения и поддержания мембранного потенциала покоя. Первой работой, положившей начало целенаправленному объяснению причин и механизмов возникновения и существования электрических явлений в живых тканях, следует считать работу В. Ю. Чаговца, выполненную в 1896 г. в лабораториях российских физиологов И.М. Сеченова и И.Р. Тарханова в Военно-медицинской академии и Санкт-Петербургском университете. В дальнейшем Ю.Бернштейном (1902) была сформулирована мембранно-ионная концепция электрогенеза живых тканей, а в 1908 г. была опубликована модель биоэлектрогенеза В. Нернста.
Результатом развития этих представлений стала мембранно-ионная теория, которую сформулировали и экспериментально обосновали в 1949-52 гг. Ходжкин, Хаксли, Катц. Согласно этой теории мембранный потенциал покоя и его изменения при возбуждении обусловлены неодинаковой концентрацией ионов натрия, калия, кальция, хлора внутри клетки и во внеклеточной жидкости, а также неодинаковой проницаемостью для этих ионов плазматической мембраны клетки. Теория, предложенная Ходжкиным и Хаксли, предлагает исчерпывающе полное биофизическое описание потенциала действия, хотя методы исследования молекулярных механизмов нервного импульса (мембранных структур, контролирующих перемещение ионов) стали доступными только в 80-х гг.
Ходжкин и Хаксли получили Нобелевскую премию по физиологии и медицине 1963 г. «за открытия, касающиеся ионных механизмов, участвующих в возбуждении и торможении в периферическом и центральном участках мембраны нервной клетки». Они разделили премию вместе с Джоном С. Эклсом.
Жидкостно-мозаичная модель биологических мембран
В 1972 г. Джонатан Сингер и Гарт Николсон предложили жидкостно-мозаичную модель, объясняющую в общих чертах организацию биологических мембран. Согласно этой модели, мембраны представляют собой двумерные растворы определенным образом ориентированных фосфолипидов и глобулярных белков которые пронизывают мембрану насквозь (интегральные белки) или погружены в ее толщу (периферические белки).

Рис. 2. Жидкостно-мозаичная модель строения плазматической мембраны.
Большая часть мембранных фосфолипидов и гликолипидов представлена в виде бислоя который играет двоякую роль. Во-первых, небольшая часть мембранных липидов специфически связана с определенными мембранными белками и, вероятно, необходима для их функционирования. Во-вторых, будучи гидрофобными, мембранные фосфолипиды являются барьером проницаемости, обеспечивая одну из базовых функций клетки – барьерную (защитную), благодаря которой внутреннее содержимое клетки надежно отделено от внешней среды.
Молекулы белков встроены (интеркалированы) в фосфолипидный матрикс клеточной мембраны. Все мембранные белки свободно перемещаются в липидном матриксе в латеральном направлении, но не могут перемещаться в поперечном направлении, т.е. от одной поверхности мембраны к другой.
Благодаря высокой химической активности, специфическими белками выполняются многие важные функции мембран связанные с распознаванием сигналов, ферментативной активностью, преобразованием энергии, переносом веществ. Белки, прикрепленные к поверхности клеточной мембраны (в основном к внутренней ее части), называют периферическими, они, как правило, являются ферментами (ацетилхолинестераза, фосфатазы, аденилатциклаза, протеинкиназы). Некоторые интегральные белки также выполняют функцию ферментов, например АТФаза. Рецепторами и антигенами мембраны могут быть как интегральные, так и периферические белки. Белки, примыкающие к мембране с внутренней стороны, являются также составной частью цитоскелета, который обеспечивает дополнительную прочность клеточной мембране и ее эластичность.
Важнейшей функцией интегральных белков является перенос веществ через клеточную мембрану. (транспортная функция) – процесс, имеющий фундаментальное значение для всех живых клеток, так как обеспечивает обмен веществ и поддержание гомеостаза. Кроме того, за счет переноса заряженных частиц – ионов - работой транспортных систем поддерживается электрическая активность плазматической мембраны, лежащая в основе раздражимости и возбудимости.
Основную роль в возникновении и поддержании электрических состояний мембраны играют следующие ее транспортные системы: первично активного транспорта – ионные насосы (помпы), - работа которых обеспечивает формирование и восстановление заряда мембраны, и вторично активного транспорта – ионные каналы, - которые играют основную роль в изменении заряда мембраны при действии раздражителя.
Насосы представляют собой белковые молекулы, обладающие свойствами переносчика и АТФ-азной активностью. Обычно указывают на существование трех ионных насосов: натрий-калиевого, кальциевого и водородного, есть основание предполагать наличие и хлорного насоса. Насосы локализуются на клеточных мембранах или мембранах органелл клеток.
В результате работы ионных насосов плазматической мембраны создаются и поддерживаются трансмембранные ионные градиенты, определяющие заряд мембраны – мембранный потенциал:
-
концентрация Na+, Ca2+, Cl– внутри клетки ниже, чем снаружи (в межклеточной жидкости)
-
концентрация K+ внутри клетки выше, чем снаружи.
Натрий-калиевый насос (Na/K-АТФаза) — это интегральный белок клеточной мембраны, обладающий, как и все другие насосы, свойствами фермента, т. е. сам переносчик обеспечивает расщепление АТФ и освобождение энергии, которую он же сам и использует. Более трети энергии АТФ, потребляемой клеткой в состоянии покоя, расходуется на перенос только ионов Na+ и К+. Этот насос имеется в мембранах всех клеток и создает характерный признак живого — градиент концентрации Na+ и К+ внутри и вне клетки, что обеспечивает формирование мембранного потенциала и вторичный транспорт веществ. Главными активаторами насоса являются гормоны (альдостерон, тироксин), ингибирует насос недостаток энергии (кислородное голодание), его специфическими блокаторами являются строфантины, особенно уабаин. Работа натриевого насоса после удаления К+ из среды сильно нарушается.
Кальциевый насос локализуется в эндоплазматическом ретикулуме и в клеточной мембране, он обеспечивает транспорт Са2+. Насос строго контролирует содержание Са2+ в клетке, поскольку изменение содержания Са2+ в ней нарушает ее функционирование. Насос переносит Са2+ либо во внеклеточную среду (в гладких мышцах), либо в цистерны ретикулума (в поперечно-полосатых мышцах) и митохондрии (внутриклеточное депо Са2+).
Протонный насос работает в митохондриях .
Хлорный насос работает, по-видимому, подобно всем другим помпам и играет главную роль в процессах торможения ЦНС.
Механизм работы ионных насосов заключается в следующем. Na+/K+-насос переносит за один цикл 2Na+ из клетки и 3К+ в клетку . Это осуществляется в результате конформации (изменения третичной структуры) молекулы белка в форму при которой его активный участок способен связывать либо Na+ (форма Е1), либо К+ (форма Е2). При конформации Е1 активная часть белка обращена внутрь клетки где и связывает Na+, вследствие чего активируется его АТФ-аза, в результате белок превращается в форму Е2 и активный участок поворачивается наружу клеточной мембраны. Теперь он теряет сродство к Na+, который отщепляется, а приобретает сродство к иону К+ и соединяется с ним. Это снова ведет к изменению конформации переносчика: форма Е2 переходит в форму Е1 , а активный участок белка опять поворачивается внутрь клетки. При этом он теряет сродство к иону К+, который отщепляется, а белок приобретает снова сродство к иону Na+, т.е. цикл повторяется.
Насос является электрогенным, поскольку за один цикл выводится из клетки два иона Na+, а возвращаются в клетку три иона К+. Энергия расходуется только на перенос Na+. На Обеспечение одного цикла работы Na/K-помпы расходуется одна молекула АТФ.
Подобным образом работают и Са-АТФазы эндоплазматического ретикулума, а также клеточной мембраны, с тем лишь различием, что переносятся только ионы Са2+ и в одном направлении: из гиалоплазмы в эндоплазматический ретикулум либо наружу клетки.
Ионные каналы – интегральные белки, которые обеспечивают пассивный транспорт ионов по градиенту концентрации. Энергией для транспорта служит разность концентрация ионов по обе стороны мембраны (трансмембранный ионный градиент), который создается работой мембранных насосов.
Наличие ионных каналов впервые было доказано для мембран нервной ткани. Структурно каналы представляет собой, как бы, «поры» которые имеют устье и селективный фильтр, а управляемые каналы — и воротный механизм. Каналы способны пропускать ионы через мембрану с огромной скоростью, через один ионный канал может проходить 107- 108 ионов в секунду. Количество каналов на единицу площади мембраны так же очень велико, поэтому суммарный заряд переносимых ионов может быть относительно большим. Движение зараженных ионов, создает ток, текущий через плазматическую мембрану клетки. Изменение величины и направления этого тока, приводит к изменению заряда самой плазматической мембраны и воспринимается клеткой как изменение окружающей среды. Этот процесс и лежит в основе электрической природы раздражимости.
Электрические и физиологические проявления возбуждения
В состоянии покоя мембрана возбудимой клетки поляризована, т.е. имеется постоянная разность потенциалов между внутренней и наружной поверхностью клеточной мембраны, причем наружная сторона заряжена положительно по отношению к внутренней. Эту разность потенциалов называют потенциалом покоя. Возникновение потенциала покоя – результат работы мембранных транспортных систем.
Ионные основы возникновения потенциала покоя. Физиологической основой потенциала покоя является неравномерное распределение различных ионов (прежде всего К+) между вне- и внутриклеточным пространством, создаваемое непрерывной работой Na+/К+ - АТФ-азы. Закачивая в клетку ионы К+, и удаляя из клетки ионы Na+, (в соотношении 3К+/2Na+), она создает мощнейший химический градиент этих ионов (10-15 раз) между вне- и внутриклеточным пространствами. Сам по себе, этот градиент не приводит к возникновению существенного заряда на мембране, т.к. заряд переносимых ионов одинаков, а электрогенность самой АТФ-азы невелика. Однако, возникающий градиент концентрации калия столь велик, что диффузия этого иона из клетки становится достаточно значимой. Кроме того, большая часть его «стравливается» через неспособные к инактивации К+ - каналы (каналы утечки).
Мембранный потенциал, возникающий в результате утечки К+, называют “равновесным калиевым потенциалом” (ЕК). Его можно рассчитать по формуле Нернста:
![]()
где R – универсальная газовая постоянная,
Т – температура (по Кельвину),
F – число Фарадея,
[К+]нар – концентрация ионов К+ снаружи клетки,
[К+]вн – концентрация ионов К+ внутри клетки.
Вынос дополнительного, положительного заряда наружу приводит к тому, что, наружная сторона плазматической мембраны заряжается положительно относительно своей внутренней стороны, т.е. возникает потенциал – потенциал покоя. Величина его близка к равновесному калиевому, но не равна ему. Эта разница объясняется тем, что свой вклад в формирование ПП вносят:
во-первых - поступление в клетку Na+ и Cl– через неселективные ионные каналы; при этом поступление в клетку Cl– дополнительно гиперполяризует мембрану, а поступление Na+ - дополнительно деполяризует ее; вклад этих ионов в формирование ПП невелик, т.к. проницаемость неселективных каналов для Cl– и Na+ в 2,5 и 25 раза ниже, чем для К+;
во-вторых – прямой электрогенный эффект Na+/К+ ионного насоса, возникающий в том случае, если ионный насос работает асимметрично (количество переносимых в клетку ионов K+ не равно количеству выносимых из клетки ионов Na+).
Поле, создаваемое избытком положительных ионов снаружи клетки формирует электрический градиент, который препятствует неограниченному выходу ионов К+ из клетки. Таким образом возникает динамическое равновесие способствующее непрерывному поддержанию определенной величины потенциала покоя.
Любая живая клетка поддерживает на мембране определенной величины потенциал. Его величина колеблется в значительных пределах, у возбудимых клеток его величина составляет обычно 60 - 90 мВ, у других тканей не превышает 10 мВ. Различные внешние воздействия, способные изменять ионную проницаемость мембраны, вызывают изменения величины ПП. Форма и последствия этих изменений зависят, при прочих равных условиях, от характеристик раздражителя.
Универсальным раздражителем для мембран возбудимых клеток является электрический ток, а все формы изменения величины ПП наблюдаются при внутриклеточном способе раздражения (с помощью электродов, один из которых введен внутрь клетки, а другой расположен на ее поверхности (см. рис.1)).
Эти формы представлены на рис 4.
При действии слабых импульсов постоянного электрического тока на мембране клетки под приложенными электродами развивается электротонический потенциал (ЭП) – сдвиг мембранного потенциала в сторону соответствующую полярности электрода. ЭП это пассивная реакция клетки на электрический раздражитель. Никакой физиологической реакцией клетки не проявляется, состояние ионных каналов и транспорт ионов при этом не изменяется, т.к. силы раздражителя не хватает для открытия воротного механизма канала, Поэтому ЭП не является возбуждением.
П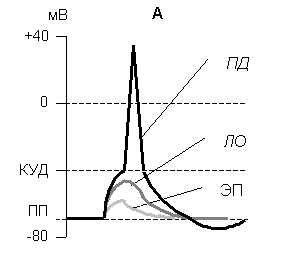 ри
действии более сильного тока возникает
и более сильный сдвиг мембранного
потенциала – локальный ответ. Локальный
ответ (ЛО) – активная реакция клетки
на электрический раздражитель, при
котором происходит открытие
потенциалчувствительных ионных каналов.
Однако, транспорт ионов при этом
изменяется незначительно, ток текущий
через открытые каналы невелик и не
способен повлиять на воротный механизм
каналов находящихся вне поля действия
электродов. Локальный ответ не проявляется
заметной физиологической реакцией
клетки, его называют местным возбуждением,
т.к. это возбуждение не распространяется
по мембранам возбудимых клеток. Такая
форма электрической активности является
основной для большинства клеток не
способных к возбуждению, и реагирующих
изменением мембранного потенциала
только в месте действия раздражителя.
ри
действии более сильного тока возникает
и более сильный сдвиг мембранного
потенциала – локальный ответ. Локальный
ответ (ЛО) – активная реакция клетки
на электрический раздражитель, при
котором происходит открытие
потенциалчувствительных ионных каналов.
Однако, транспорт ионов при этом
изменяется незначительно, ток текущий
через открытые каналы невелик и не
способен повлиять на воротный механизм
каналов находящихся вне поля действия
электродов. Локальный ответ не проявляется
заметной физиологической реакцией
клетки, его называют местным возбуждением,
т.к. это возбуждение не распространяется
по мембранам возбудимых клеток. Такая
форма электрической активности является
основной для большинства клеток не
способных к возбуждению, и реагирующих
изменением мембранного потенциала
только в месте действия раздражителя.
Рис. 4. Изменение мембранного потенциала при действии электрического тока разной силы
ЭП - электротонический потенциал,
ЛО – локальный ответ,
ПД – потенциал действия.
При достижении силы раздражителя определенной (пороговой) величины происходит быстрое, активное изменение потенциала покоя - потенциал действия (ПД), - быстрое колебание (спайк) мембранного потенциала, возникающее при возбуждении нервных, мышечных, некоторых железистых и растительных клеток; электрический сигнал, обеспечивающий быструю передачу информации в организме. Потенциал действия лежит в основе процесса возбуждения. Он характеризуется тем, что значение потенциала покоя клетки очень быстро уменьшается до 0 (деполяризация), и даже приобретает положительное значение (+20…+30 мВ), т.е. внутренняя сторона мембраны заряжается положительно относительно наружной. Затем значение МП быстро возвращается к исходному уровню. Сильная деполяризация клеточной мембраны во время ПД приводит к развитию физиологических проявлений возбуждения (сокращение, секреция и др.). Потенциал действия называют распространяющимся возбуждением, поскольку, возникнув в одном участке мембраны, он быстро распространяется во все стороны.
Механизм развития ПД практически одинаков для всех возбудимых клеток.
Ионные основы возникновения потенциала действия. В основе потенциала действия лежит увеличение проницаемости плазматической мембраны, прежде всего для ионов Na+, приводящее к нарушению (изменению) распределения этого иона между вне- и внутриклеточным пространством. Это изменение проницаемости вызывается действием раздражителя, который либо уменьшает заряд мембраны (деполяризует ее), что приводит к открытию воротного механизма электроуправляемых Na+ - каналов у места действия раздражителя, либо действует на воротный механизм хемо-управляемых Na+ - каналов через соответствующий рецептор.
Благодаря наличию химического градиента, через открытые каналы ионы Na+ свободно поступают в клетку и продолжают деполяризовать ее мембрану у места действия раздражителя. Когда величина деполяризации достигнет критической величины (порога), достаточной для открытия воротного механизма соседних каналов, процесс становится необратимым и распространяющимся.
Это состояние автоматического прогрессирующего нарушения мембранного заряда и есть суть возбуждения. Возбуждение пропорционально величине деполяризации, т.е. количеству входящих в клетку ионов, а величина деполяризации характеризуется амплитудой ПД и зависит от силы раздражителя.
Сильная и длительная деполяризация мембраны ведет к инактивации натриевых каналов и прекращению входа Na+. Недостаток положительно заряженных ионов (Na+) снаружи мембраны нарушает электрохимическое равновесие - потенциал покоя, что приводит к повышению калиевой проницаемости. Выход ионов калия наружу через неинактивируемые К+ каналы и потенциалчувствительные, управляемые К-каналы продолжается до восстановления электрохимического градиента т.е. потенциала покоя. Это фаза процесса возбуждения называется фазой реполяризацией.
Окончание процесса возбуждения и переход клетки в состояние физиологического покоя сопровождается активацией натрий - калиевого насоса перекачивающего ионы Nа+ из клетки, а ионы К+ - внутрь ее. В клетке восстанавливается способность к следующему акту возбуждения.
Таким образом, сущность процесса возбуждения заключается в следующем. Все клетки организма имеют электрический заряд, обеспечиваемый неодинаковой концентрацией анионов и катионов внутри и вне клетки. Различная концентрация анионов и катионов внутри и вне клетки является следствием работы ионных насосов и неодинаковой проницаемости клеточной мембраны для разных ионов. При действии раздражителя на клетку возбудимой ткани изменяется проницаемость ее мембраны (сначала повышается для Na+ и быстро возвращается к норме, затем также, но более медленно изменяется для К+), вследствие чего ионы быстро перемещаются в клетку и из клетки согласно электрохимическому градиенту. Эта ответная реакция возбудимой клетки на раздражение, выражающаяся в быстром перемещении ионов в клетку и из клетки согласно электрохимическому градиенту, и есть возбуждение, основой которого является потенциал покоя.
Одиночный цикл возбуждения.
К возбудимым тканям относятся только те, клетки которых генерируют потенциал действия (ПД). Это мышечные и нервные клетки. Нередко к возбудимым тканям необоснованно относят и «железистую ткань», хотя железистой ткани нет, а имеются различные железы и железистый эпителий как вид тканей. В процессе активной деятельности железы в ней действительно регистрируются биоэлектрические явления, поскольку железа как орган состоит из различных клеток: соединительной ткани, эпителиальной, мышечной. ПД проводится по мембранам нервных и мышечных клеток, с его помощью передается информация и обеспечивается управление деятельностью клеток организма.
Невозбудимыми тканями являются эпителиальная и соединительная (собственно соединительная, ретикулярная, жировая, хрящевая, костная и гемотопоэтические ткани в совокупности с кровью), клетки этих тканей хотя и способны изменять свой мембранный потенциал, но не генерируют ПД при действии на них раздражителя.
Основными физиологическими свойствами возбудимых тканей являются: возбудимость, проводимость, рефрактерность, лабильность. Специфическим свойством мышечной ткани является сократимость.
Возбудимость — это свойство некоторых тканей генерировать потенциал действия (ПД) в ответ на раздражение. Развитие ПД возможно только при действии раздражителей, которые вызывают деполяризацию клеточной мембраны. Раздражители вызывающие гиперполяризацию мембран будут приводить к процессу обратному возбуждению – торможению.
Возбудимость может быть охарактеризована кривой потенциала действия, в которой выделяют несколько фаз (рис.1 А). Отметим, что единой терминологии в классификации этих фаз нет, поэтому будем использовать наиболее часто употребляемые названия.
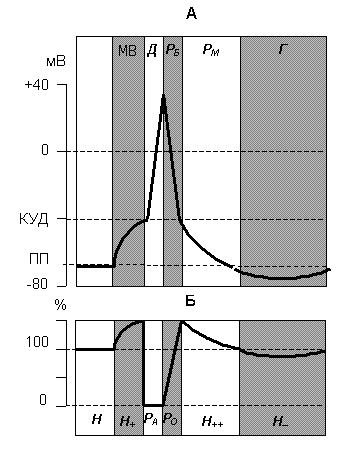
Рис. 1. Изменение мембранного потенциала (А) и возбудимости клетки (Б) в разные фазы потенциала действия.
МВ – фаза местного возбуждения;
Д – фаза деполяризации;
РБ – фаза быстрой реполяризации;
РМ – фаза медленной реполяризации;
Г – фаза следовой гиперполяризации;
Н – период нормальной возбудимости;
РА – период абсолютной рефрактерности;
РО – период относительной рефрактерности;
Н+ – период первичной экзальтации;
Н++ – период экзальтации;
Н– – период субнормальной возбудимости.
Вначале, под действием раздражителя, развивается местное возбуждение (фаза начальной деполяризации) - процесс медленной деполяризации мембраны от мембранного потенциала до критического уровня деполяризации (КУД). Если этот уровень не будет достигнут – ПД не формируется, а развивается только локальный ответ.
Разность между мембранным потенциалом покоя и критическим уровнем деполяризации называют пороговым потенциалом, его величина определяет возбудимость клетки – чем больше пороговый потенциал, тем меньше возбудимость клетки.
Время фазы начальной деполяризации очень короткое, на кривой ПД она регистрируется только при большой развертке, и чаще всего является составной частью общей фазы деполяризации. Эта фаза развивается при достижении КУД, за счет открытия всех потенциалчувствительных Na+ - каналов и лавинообразного входа ионов Na+ в клетку по градиенту концентрации (входящий натриевый ток). В результате, мембранный потенциал очень быстро уменьшается до 0, и даже приобретает положительное значение. Графически – это восходящая часть кривой потенциала действия. В результате инактивации Na+ - каналов и прекращения поступления Na+ в клетку, рост кривой ПД прекращается и начинается ее снижение. Явление изменения знака мембранного потенциала называют реверсией заряда мембраны.
По мнению некоторых исследователей, фаза деполяризации заканчивается уже тогда, когда мембранный потенциал становится равным нулю, и весь период, когда величина мембранного потенциала превышает величину 0 мВ, следует считать отдельной фазой реверсии, т.к. ионные токи, определяющие развитие этой части ПД, имеют характерные особенности.
Период времени, в течение которого мембранный потенциал имеет положительное значение, называется овершут.
Нисходящая част кривой ПД – фаза реполяризации. Она определяется выходящим калиевым током. Калий выходит через постоянно открытые каналы утечки, ток через которые резко возрастает из-за изменения электрического градиента вызванного нехваткой снаружи ионов Na+ и через потенциалочувствительные, управляемые К+- каналы, которые активируются на пике ПД.
Различают быструю и медленную реполяризацию. В начале фазы, когда активны оба типа каналов, реполяризация происходит быстро, к концу фазы, ворота потенциалочувствительных К+- каналов закрываются, интенсивность калиевого тока снижается и реполяризация замедляется. Она прекращается тогда, когда положительный заряд снаружи мембраны вырастет настолько, что окончательно затруднит выход калия из клетки.
Фазу медленной реполяризации называют иногда отрицательным следовым потенциалом, что не совсем верно, так как эта фаза не является потенциалом по определению и не является следовым процессом по механизму.
Фаза следовой гиперполяризации (следовой положительный потенциал) – увеличение мембранного потенциала выше величины потенциала покоя, которое наблюдается у нейронов. Развивается за счет остаточного калиевого тока и за счет прямого электрогенного эффекта активировавшейся Na+/K+ АТФ-азы.
Механизм наблюдаемой иногда следовой деполяризации (следовой отрицательный потенциал) до конца не ясен.
Изменение возбудимости клетки при развитии возбуждения. Рефрактерность.
Возбудимость в различные фазы развития одного цикла возбуждения, вообще является переменной величиной. В ходе развития одного цикла возбуждения возбудимость изменяется в сторону, как повышения, так и понижения. Повышение возбудимости называется экзальтацией, понижение – рефрактерностью.
В изменении возбудимости от момента нанесения раздражения до завершения одиночного цикла возбуждения отмечается несколько периодов (фаз). (Рис.1. Б)
В период развития местного возбуждения наблюдается некоторое повышение возбудимости, которое получило название первичной экзальтации. Каждое нанесенное в это время дополнительное раздражение, по силе даже ниже порогового, ускоряет развитие местного потенциала. Это связано с тем, что пороговый потенциал уменьшается, и открытие воротного механизма Na+ -каналов облегчается.
Как только местное возбуждение достигает критической величины и переходит в потенциал действия (фаза деполяризации), возбудимость начинает быстро снижаться и в точке пика потенциала практически становится равной нулю. Это связано с полной инактивацией Na+ -каналов на пике ПД.
Время, в течение которого происходит это снижение возбудимости называется абсолютной рефрактерной фазой (периодом), а само снижение возбудимости - абсолютной рефрактерностью. Раздражение любой сверхпороговой силы, нанесенное в этот период, практически не может повлиять на развитие текущего возбуждения (потенциала действия).
В фазе реполяризации возбудимость мембраны последовательно восстанавливается, до исходного уровня, за счет постепенного восстановления активности инактивированных Na+ -каналов. Пока активны не все каналы – то этот период называется относительной рефрактерной фазой, а состояние, в котором находится живой объект - относительной рефрактерностью. Эта фаза продолжается до восстановления заряда мембраны до величины, соответствующей критическому уровню деполяризации. Раздражение, нанесенное в этот период, может вызвать усиление возбуждения только в том случае, если по силе оно будет больше величины порогового потенциала Длительность относительной рефрактерной фазы может быть значительно больше, чем абсолютной.
Вслед за периодом относительной рефрактерности наступает фаза экзальтации (повышенной возбудимости). Это связано с тем что мембранный потенциал снижается до величины КУД, при которой восстанавливается активность большей части Na+ каналов, а разница между величиной мембранного потенциала и КУД – пороговый потенциал – минимальна. В этой фазе может возникнуть повторная волна возбуждения даже на раздражения, которые значительно ниже порогового потенциала. Фаза экзальтации длится до тех пор, пока не восстановится исходная величина мембранного потенциала – потенциал покоя, при этом восстанавливается исходная величина возбудимости.
В фазы следовой гипер- и деполяризации возбудимость меняется незначительно и связана с колебаниями порогового потенциала.
Биологический смысл фазового изменения возбудимости в ходе развития одиночной волны возбуждения заключается в следующем.
Начальная фаза повышения возбудимости обеспечивает условие, при котором каждый дополнительный раздражитель ускоряет процесс подготовки (местное возбуждение) к специфической (для данной ткани) приспособительной реакции.
Состояние абсолютной рефрактерности позволяет данной ткани «без помех» осуществлять текущую приспособительную реакцию. Если бы в этих условиях возбудимость была нормальной, то дополнительное раздражение, вызвав дополнительное возбуждение, могло бы исказить эту реакцию, превратив ее в избыточную или недостаточную для данных условий.
Абсолютная рефрактерность защищает ткань от чрезмерных энергетических трат в процессе осуществления текущей приспособительной реакции. Сходную роль играет и относительная рефрактерность, с той разницей, что в данном случае живое образование в состоянии реагировать на раздражения, требующие срочного ответа. Именно поэтому для большинства тканей и органов, работающих непрерывно и не имеющих длительных периодов физиологического покоя (например, сердце), характерна более длительная по сравнению со скелетной мускулатурой рефрактерность.
Кроме того, рефрактерность - один из факторов, определяющих максимальный (предельный) ритм импульсации клетки, что лежит в основе например кодирования и декодирования сигнала структурами нервной системы, регуляции восприятия, сокращения, обеспечении одностороннего проведения возбуждения по нервам и др.
Состояние зкзальтации создает условия готовности ткани к ответу на повторное раздражение не только прежней силы, но и более слабой.
Лабильность, или функциональная подвижность, одно из физиологических свойств живых тканей. Это свойство описано в 1892 г. Н. Е. Введенским, который установил, что скорость протекания процесса возбуждения в тканях различна. Каждая возбудимая ткань способна на раздражение отвечать только определенным количеством волн возбуждения. Так, нервное волокно способно воспроизводить до 1000 импульсов в секунду, поперечно-полосатая мышца только 200 250 имп/с.
Мерой лабильности, по Н. Е. Введенскому, является то наибольшее количество волн возбуждения, которое возбудимая ткань может воспроизводить в 1 с в точном соответствии с ритмом наносимых раздражений без явлений трансформации (переделки) ритма, т.е. не уменьшая и не увеличивая его.
Лабильность величина подвижная и может изменяться в достаточно широких пределах. В частности, лабильность широко варьирует в процессе ритмического раздражения. В одних случаях вследствие взаимодействия волн возбуждения лабильность может повыситься, в других понизиться. Повышение лабильности может привести к тому, что недоступные ранее ритмы деятельности станут доступными. На основании этого А. А. Ухтомский сформировал представление об «усвоении ритма», как способности ткани отвечать на раздражение более высоким или более низким ритмом возбуждения по сравнению с его исходным уровнем. Усвоение ритма зависит от текущих изменений обмена веществ в ткани во время ее деятельности
Явление усвоения ритма играет важную роль в процессах врабатывания и тренировки. Снижение лабильности, происходящее в процессе деятельности, приводит к иному результату, способность ткани к ритмической работе уменьшается. Лабильность может быть измерена косвенным путем по величине хронаксии (см. ниже) возбудимых тканей. Чем короче хронаксия, тем выше лабильность. Определение лабильности весьма важно в физиологии труда и спорта.
Проводимость - способность живой ткани проводить возбуждение, которое, возникая в рецепторе, распространяется по нервной системе и является для организма информацией, закодированной в нейроне в виде электрических или химических сигналов. Способностью к проведению возбуждения обладают практически все возбудимые ткани, но наиболее ярко она выражена в нервной ткани, для которой проводимость является одной из функций.
Подробно механизм и закономерности распространения возбуждения по мембранам возбудимых клеток рассмотрен в отдельном занятии.
Законы раздражения.
Процесс возбуждения начинается с действия на возбудимую клетку какого либо раздражителя.
Раздражитель - любое изменение внешней или внутренней среды организма, воспринимаемое клетками и вызывающее ответную реакцию. По своей природе раздражители делят физические (электрические, механические, температурные, световые) и химические.
В зависимости от степени чувствительности клеток к тому или иному раздражителю их подразделяют на адекватные и неадекватные. Адекватный раздражитель — это такой раздражитель, к которому клетка обладает наибольшей чувствительностью вследствие наличия специальных структур, воспринимающих этот раздражитель. Так, адекватным раздражителем для фоторецепторов сетчатки глаза, например, являются световые волны, адекватным раздражителем нейронов являются медиаторы и электрические импульсы.
Неадекватные раздражители в естественных условиях существования организма не воздействуют на возбудимые структуры. Однако, при достаточной силе и продолжительности действия, могут вызвать ответную реакцию со стороны возбудимых тканей, например, удар в глаз при достаточной силе может вызвать ощущение вспышки света.
В условиях физиологического эксперимента в качестве раздражителя чаше всего используют электрический ток. Электрический ток легко дозировать, и он является адекватным раздражителем для возбудимых тканей, так как их функциональная активность всегда сопровождается электрическими явлениями.
Определенную зависимость между действием раздражителя и ответной реакцией возбудимой ткани отражают законы раздражения. К законам раздражения относятся:
Закон силы.
Для возникновения возбуждения решающее значение имеет сила раздражителя. Возбуждение возникают только в том случае, если сила действующего раздражителя достигает минимальной, критической величины, которая характеризуется порогом возбуждения. По отношению к этой величине, по своей силе раздражители могут быть подпороговыми, пороговыми и надпороговыми.
Подпороговый раздражитель - это раздражитель такой силы, который не вызывает видимых изменений, но обусловливает возникновение физико-химических сдвигов в возбудимых тканях, например локального ответа. Однако степень этих сдвигов недостаточна для возникновения распространяющегося возбуждения.
Пороговый раздражитель - это раздражитель минимальной силы, который впервые вызывает минимальную измеримую ответную реакцию со стороны возбудимой ткани. Именно эту пороговую силу раздражителя называют порогом раздражения или возбуждения. Порог раздражения и является мерой возбудимости ткани. Между порогом раздражения и возбудимостью существует обратная зависимость: чем выше порог раздражения, тем ниже возбудимость, чем ниже порог раздражения, тем возбудимость выше. При достижении раздражителем величины порога, возникновение потенциала действия становится неизбежным.
Следует отметить, что порог раздражения показатель достаточно изменчивый и значительно зависит от исходного функционального состояния возбудимой ткани и практически никак не зависит от характеристик самого раздражителя
Надпороговый раздражитель - это раздражитель, сила которого выше, чем сила порогового раздражителя.
Закон силы - характеризует взаимосвязь между силой раздражителя и электрическим ответом, он может быть применен для простых и сложных систем.
Простая возбудимая система – это одна возбудимая клетка, которая реагирует на раздражитель как единое целое. Исключением является сердечная мышца, которая вся реагирует как одна клетка. Закон силы для простых возбудимых систем - подпороговые раздражители не вызывают возбуждения, а пороговые и сверхпороговые раздражители вызывают сразу максимальное возбуждение (Рис. 2).
При подпороговых значениях раздражающего тока возбуждение (электротонический потенциал, локальный ответ) носит местный (не распространяется), градуальный (сила реакции пропорциональная силе действующего стимула) характер. При достижении порога возбуждения возникает ответ максимальной силы (ПД). Амплитуда ответа (амплитуда ПД) не изменяется при дальнейшем увеличении силы раздражителя.
Закон силы для простых возбудимых систем известен как закон «все ли ничего».
Сложная возбудимая система – система, состоящая из множества возбудимых элементов (мышца включает множество двигательных единиц, нерв – множество аксонов). Отдельные элементы (клетки) системы имеют неодинаковые пороги возбуждения.
Закон силы для сложных возбудимых систем - амплитуда ответа пропорциональна силе действующего раздражителя (при значениях силы раздражителя от порога возбуждения самого легковозбудимого элемента до порога возбуждения самого трудновозбудимого элемента) (рис. 3). Амплитуда ответа системы пропорциональна количеству вовлеченных в ответ возбудимых элементов. При возрастании силы раздражителя в реакцию вовлекается все большее число возбудимых элементов.
В случае сложных систем, от силы раздражителя будет зависеть не только электрический, но и физиологический (функциональный) ответ ткани, например сила сокращения. В этом случае закон силы будет звучать следующим образом: чем больше сила раздражителя, тем выше, до определенного предела, ответная реакция со стороны возбудимой ткани. Этот предел будет определяться функциональными возможностями ткани.
Ответ минимальной силы – едва заметное сокращение - возникнет при достижении раздражителем пороговой величины. При этом сократятся мышечные волокна имеющие наименьший порог возбуждения.
Ответная реакция на надпороговый раздражитель будет выше и по мере его увеличения некоторое время также возрастает за счет вовлечения в сокращение все новых мышечных волокон, которые имеют более высокие пороги возбуждения. По достижении определенной величины раздражителя, рост силы сокращения прекратится, значит, в сокращение вовлечены все мышечные волокна. Такую ответную реакцию называют максимальной, а степени силы раздражителя, находящиеся между пороговой и максимальной – субмаксимольными.
|
Амплитуда ответа
а лЛ И Т У Д а ответа
|
|
Амплитуда ответа
|
|
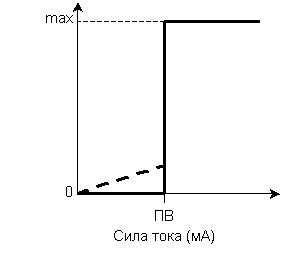
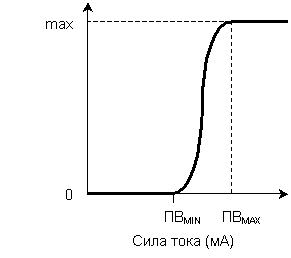
Рис. 2. Зависимость силы реакции простой Рис. 3. Зависимость силы реакции сложной
возбудимой системы от силы раздражителя. возбудимой системы от силы раздражителя. ПВ – порог возбуждения. ПВMIN – порог возбуждения самого
легковозбудимого элемента,
ПВMАХ – порог возбуждения самого
трудновозбудимого элемента.
Увеличение силы раздражителя выше максимального какое-то время не сказывается на величине ответной реакции. Такую силу раздражителя называют супермаксимальной. Но при достаточно большом увеличении силы раздражителя, сила ответной реакции начинает снижаться. Такую величину силы раздражителя называют пессимальной.
Пессимальный ответ и есть тот определенный предел, до которого может расти ответная реакция. Превышение этого предела при спортивных, интеллектуальных, эмоциональных и любых других нагрузках не имеет никакого физиологического смысла для получения результата.
Закон силы-времени (силы-длительности)
Эффективность раздражителя зависит не только от силы, но и от времени его действия. Длительность действия раздражителя, способна компенсировать недостаток силы раздражителя и при его недостатке привести, тем не менее, к возникновению распространяющегося потенциала действия, поэтому важно определять не только пороговую силу, но пороговую длительность раздражителя. Учение о хронаксии как пороговом времени необходимом для возникновения возбуждения было создано французским ученым Лапиком.
Связь между силой и временем действия раздражителя характеризует закон силы длительности - сила раздражителя, вызывающего процесс распространяющегося возбуждения, находится в обратной зависимости от длительности его действия, т.е., чем больше сила раздражителя, тем меньше времени он должен действовать для возникновения возбуждения. Зависимость между силой раздражителя и продолжительностью его воздействия, необходимого для возникновения минимальной ответной реакции живой структуры, очень хорошо можно проследить на так называемой кривой силы - времени (кривая Гоорвега - Вейса - Лапика) (Рис 4).
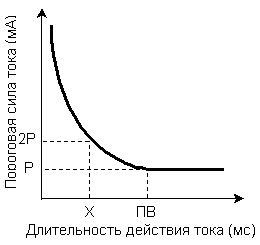
Рис.4.Зависимость пороговой силы раздражителя от времени его действия. Р – реобаза, ПВ – полезное время, Х – хронаксия.
Из кривой следует, что ток ниже некоторой минимальной величины не вызывает возбуждение, как бы длительно он не действовал, и как бы ни велика была сила раздражителя, при недостаточной длительности его воздействия ответной реакции не будет.
Минимальная сила раздражителя, способная, при неограниченном времени действия вызвать возбуждение, была названа Лапиком реобазой. Наименьшая длительность действия раздражителя силой в одну реобазу, достаточная для возникновения ответной реакции называется – полезным временем.
Но определение точки соответствующей на кривой величине полезного времени по техническим причинам затруднено, поэтому Лапиком было предложено измерять не полезное время, а условную величину – хронаксию.
Хронаксия – это наименьший промежуток времени, в течение которого ток, равный по силе удвоенной реобазе, вызывает в ткани возбуждение. Она и есть показатель пороговой длительности раздражения. Хронаксия измеряется в (тысячные доли секунды). По величине хронаксии можно судить о скорости возникновения возбуждения в ткани: чем меньше хронаксия, тем быстрее возникает возбуждение.
Определение хронаксии - хронаксиметрия - получило широкое распространение в физиологии спорта, в клинике, для определения функциональных возможностей и сохранности возбудимых тканей.
Закон крутизны нарастания силы раздражителя (закон аккомодации, закон Дюбуа-Реймона)
Для возникновения возбуждения имеет значение не только сила и время действия тока, но и скорость нарастания силы раздражителя в единицу времени от нулевой величины до величины порога. Степень крутизны, т.е. величина прироста силы раздражения в единицу времени, представляет собой градиент раздражения. Зависимость ответной реакции от градиента раздражения характеризует закон градиента (крутизны нарастания) раздражения - чем выше градиент раздражения, тем сильнее (до определенных пределов) ответная реакция возбудимого образования (рис 5).
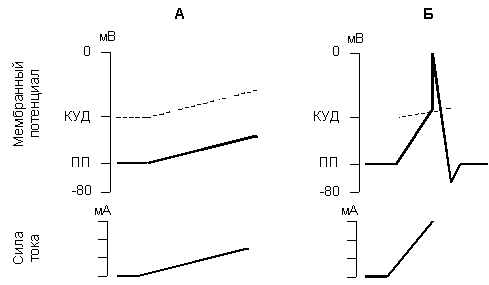
Рис. 5. Изменение мембранного потенциала и критического уровня деполяризации при медленном (А) и быстром (Б) нарастании силы раздражающего тока.
При действии медленно нарастающего раздражителя возбуждение возникает при его гораздо большей силе, так как происходит приспосабливание возбудимой ткани к действию этого раздражителя, что получило название аккомодации. Аккомодация обусловлена тем, что при действии медленно нарастающего раздражителя в мембране возбудимой ткани происходит повышение критического уровня деполяризации. При снижении скорости нарастания силы раздражителя до некоторого минимального значения потенциал действия вообще не возникает.
Причина заключается в том, что деполяризация мембраны является пусковым стимулом к началу двух процессов: быстрого, ведущего к повышению натриевой проницаемости, и тем самым обусловливающего возникновение потенциала действия, и медленного, приводящего к инактивации натриевой проницаемости и как следствие этого - окончанию потенциала действия. При быстром нарастании стимула повышение натриевой проницаемости успевает достичь значительной величины прежде, чем наступит инактивация натриевой проницаемости. При медленном нарастании тока на первый план выступают процессы инактивации, приводящие к повышению порога или ликвидации возможности генерировать ПД вообще.
Способность к аккомодации различных структур неодинакова. Наиболее высокая она у двигательных нервных волокон, а наиболее низкая у сердечной мышцы, гладких мышц кишечника, желудка.
Базисные знания студентов, необходимые ля реализации целей занятия.
-
Биологический смысл раздражимости. Физиологические основы процесса возбуждения.
-
История наблюдения биоэлектрических явлений.
-
Современные представления о строении биологических мембран. Роль мембранных транспортных систем в обеспечении электрической активности клетки.
-
Электрические и физиологические проявления возбуждения. Ионные основы возникновения потенциала покоя. Изменения электрического состояния клетки при возбуждении. Потенциал действия.
-
Одиночный цикл возбуждения и его фазы. Изменение возбудимости клетки при развитии возбуждения. Рефрактерность.
-
Лабильность, ее физиологический смысл и значение.
-
Законы раздражения; сила и длительность раздражителя, градиент раздражения.
-
Новая кора большого мозга. Структурно-функциональная характеристика.
Вопросы для самоподготовки к занятию.
1. Понятие о возбудимых тканях. Общие свойства возбудимых тканей.
2. Что такое возбуждение?
3. Что такое раздражитель и какие виды раздражителей существуют?
4. Основные положения мембранно-ионной теории возникновения биоэлектрических потенциалов.
5. Какова причина возникновения мембранного потенциала? Потенциал покоя. Какие ионы принимают участие в их поддержании?
6. Понятие о критическом уровне деполяризации.
7. Что называют деполяризацией?
8. Потенциал действия. Ионный механизм возникновения потенциала действия.
9. Как изменяется возбудимость в различные фазы ПД?
10. Смысл закона “всё или ничего”?
11. Что такое гиперполяризация?
Выполните следующие лабораторные работы
1. Приготовление нервно-мышечного препарата лягушки (см. Практикум по Нормальной физиологии/ Под ред. Н.А. Агаджаняна. - м., 1996.-с. 183-184.)
Необходимы: лягушка, препаровальная дощечка, препаровальный набор,
марлевые салфетки, раствор Рингера.
Цель работы: освоить методику приготовления нервно-мышечного препарата.
Ход работы
1. Обездвиживают лягушку декапитацией и разрушением спинного мозга. Для этого вводят браншу ножниц в полость рта, одним движением производят декапитацию. В обнажившийся разрез центральной нервной системы вводят разрушитель и разрушают спинной мозг. Качество разрушения центральной нервной системы контролируют по отсутствию тонуса конечностей.
2. Берут лягушку за задние лапки спинкой вверх. Перерезают позвоночник примерно посредине туловища. Отвисшую вниз верхнюю часть туловища и внутренности убирают, сделав разрезы справа и слева сверху вниз вдоль позвоночника до лонного сочленения, и, захватив одной рукой позвоночник, а другой край кожи, снимают кожу с обеих лапок.
3. Берут препарат за остаток позвоночника так, чтобы задние лапки свисали вниз и вырезают копчиковую кость - уростиль. При этом бранши ножниц проводят вдоль копчиковой кости так, чтобы не задеть нервные стволики. Разрезают продольным разрезом позвоночник и лонное сочленение. Из получившихся двух препаратов задних лапок лягушки один помещают в раствор Рингера, другой используют для дальнейшей препаровки.
4. Располагают бедро задней поверхностью вверх. Препаровальными крючками раздвигают мышцы и обнажают седалищный нерв. Взявшись за остаток позвоночника, выделяют нервный пучок, в составе которого идет седалищный нерв, а затем и сам седалищный нерв.
5. Вылущивают головку бедра из тазобедренного сустава и убирают мышцы бедра. В результате этих операций получается препарат реоскопической лапки.
6. Для приготовления нервно-мышечного препарата пересекают ахилово сухожилие и, взяв за него, отделяют икроножную мышцу до коленного сустава. Оставшиеся кости и мышцы голени убирают. В результате получается нервно-мышечный препарат, состоящий из седалищного нерва, икроножной мышцы и бедренной косточки.
2. Воспроизведение опытов Гальвани (см. Практикум с. 192-193).
Необxодимы: лягушка, препаровальная дощечка, препаровальный набор,
марлевые салфетки, раствор Рингера, вилочка Гальвани.
Цель работы: доказать наличие потенциала покоя в икроножной мышце лягушки.
Ход работы
1. Готовят реоскопическую лапку.
2. Воспроизводят первый опыт Гальвани. Для этого прикасаются вилочкой Гальвани, состоящей из медной и цинковой пластинок, к нерву. Наблюдают ответную реакцию.
3. Воспроизводят второй опыт Гальвани. Для этого делают разрез на икроножной мышце и осторожно набрасывают нерв на мышцу так, чтобы он попал одновременно на поврежденный и неповрежденный участки мышцы. Наблюдают ответную реакцию.
4. В отчетах зарисовывают схему первого и второго опытов Гальвани.
4. Опыт Маттеучи (вторичный тетанус) (см. Практикум с. 193).
Необходимы: лягушка, препаровальная дощечка, препаровальный набор,
марлевые салфетки, раствор Рингера, электорстимулятор.
Цель работы: доказать наличие потенциала действия в икроножной мышце.