

Продолжительность изучения темы_______________ часов
Из них на занятие ___________ часов; самостоятельная работа_________ часов.
Место проведения учебная комната
Цель: Знать рефлекторный принцип деятельности центральной нервной системы; Знать особенности и принципы проведения возбуждения в ЦНС; Уметь выявить и оценить состояние центральных и периферических звеньев рефлекторной дуги.
Задачи:
Знать основные структурные звенья рефлекторной дуги и их функциональное значение;
Знать основные механизмы переработки информации в нервных центрах на основе электрических, химических механизмов передачи возбуждения;
Знать основные медиаторы, передающие возбуждение в синапсах ЦНС;
Знать основные принципы распространения возбуждения в ЦНС (иррадиация, конвкргениция, пролонгирование) и их значение для организма;
Знать свойства нервных центров (одностороннее проведение, задержка, суммация, трансформация, посттетаническая потенция утомление, тонус и др.)
Уметь оценить роль и функциональное состояние различных звеньев рефлекторной дуги в норме и при нарушении их целостности.
Методические рекомендации по самоподготовке.
Появление многоклеточных организмов явилось начальным стимулом для дифференциации клеток и специализации части этих клеток в системы связи, что привело в конечном итоге к формированию сложнейшей нервной системы млекопитающих и человека. Нервная системарегулирует деятельность всех органов и систем, обусловливая их функциональное единство, и обеспечивает связь организма как единого целого с внешней средой.
Нервная система условно подразделяется на два больших отдела - соматическую, или анимальную, нервную систему ивегетативную, или автономную, нервную систему.
Соматическая нервная система осуществляет преимущественно функции связи организма с внешней средой, обеспечивая чувствительность и движение вызывая сокращение скелетной мускулатуры. Так как функции движения и чувствования свойственны животным и отличают их от растений, эта часть нервной системы получила название анимальной (животной).
Вегетативная нервная система оказывает влияние на процессы так называемой растительной жизни, общие для животных и растений (обмен веществ, дыхание, выделение и др.), отчего и происходит ее название (вегетативная - растительная). Обе системы тесно связаны между собой, однако вегетативная нервная система обладает некоторой долей самостоятельности и не зависит от нашей воли, вследствие чего ее также называют автономной нервной системой. Ее делят на две части симпатическую ипарасимпатичесакую.
В нервной системе выделяют центральную часть - головной и спинной мозг - центральная нервная система и переферическую, представленную отходящими от головного и спинного мозга нервами, - переферическая нервная система. На разрезе мозга видно, что он состоит из серого и белого вещества.
Серое веществообразуется скоплениями нервных клеток (с начальными отделами отходящих от их тел отростков). Отдельные ограниченные скопления серого вещества носят названия ядер.
Белое веществообразуют нервные волокна, покрытые миелиновой оболочкой (отростки нервных клеток, образующих серое вещество). Нервные волокна в головном и спинном мозге образуют проводящие пути
Переферические нервы в зависимости от того, из каких волокон (чувствительных либо двигательных) они состоят, подразделяются на чувствительные, двигательные и смешанные. Тела нейронов, отростки которых составляют чувствительные нервы, лежат в нервных узлах вне мозга. Тела двигательных нейронов лежат в передних рогах спинного мозга или двигательных ядрах головного мозга.
Центральная нервная система(ЦНС) - часть нервной системы, включающая головной и спинной мозг, которые выполняют в организме человека и животных ряд сложнейших функции.
Деятельность мозга направленную на выполнение этих функций можно разделить на пять основных категорий:
ощущение- возникающее в нервной системе в результате восприятия органами чувств изменений внешней среды;
движение- изменения состояния мышц организма возникающее под действием сигналов нервной системы;
внутренняя регуляция - регулирование работы внутренних органов в зависимости от состояния внешней или внутренней среды;
регуляция продолжения рода– контроль гормональной регуляции репродуктивных функций организма, а также регуляция полового поведения;
адаптация- обеспечение приспособления организма к изменяющимся условиям внешней среды.
И.П. Павлов показал, что центральная нервная система может оказывать три рода воздействий на органы:
- пусковое, вызывающее либо прекращающее функцию органа (сокращение мышцы, секрецию железы);
- сосудодвигательное, изменяющее ширину просвета сосудов и тем самым регулирующее приток к органу крови;
- трофическое, повышающее или понижающее обмен веществ и, следовательно потребление питательных веществ и кислорода. Благодаря этому постоянно согласуется функциональное состояние органа и его потребность в питательных веществах и кислороде. Когда к работающей скелетной мышце по двигательным волокнам направляются импульсы, вызывающие ее сокращение, то одновременно по вегетативным нервным волокнам поступают импульсы, расширяющие сосуды и усиливающие обмен веществ. Тем самым обеспечивается энергетическая возможность выполнения мышечной работы.
Центральная нервная система воспринимает афферентную (чувствительную) информацию, возникающую при раздражении специфических рецепторов, и в ответ на это формирует соответствующиеэфферентныеимпульсы, вызывающие изменения в деятельности определенных органов и систем организма.
Анализ функций ЦНС позволяет сформулировать значение центральной нервной системы:
Центральная нервная система обеспечивает взаимную связь отдельных органов и систем, согласует и объединяет их функции. Благодаря этому организм работает как единое целое. Точность контроля за работой внутренних органов достигается существованием двусторонней круговой связи между центральной нервной системой и периферическими органами.
Центральная нервная система осуществляет взаимодействие организма, как единого целого,с внешней средой,а также индивидуальное приспособление к внешней среде – поведение. Этот вид деятельности основанный на врожденных механизмах называют низшей нервной деятельностью (инстинкты), а на приобретенных - высшей нервной деятельностью (условные рефлексы).
Головной мозг является органом психической деятельности.В результате поступления нервных импульсов в клетки коры головного мозга возникают ощущения и на их основе проявляются специфические качества высокоорганизованной материи - процессы сознания и мышления. Психическая деятельность – это идеальная, субъективно осознаваемая деятельность организма осуществляемая с помощью нейрофизиологических процессов. То есть психическая деятельность реализуется с помощью ВНД, а не является ею.
Рефлекторная теория и принципы рефлекторной деятельности. Рефлекс.
Рефлекс – основная форма нервной деятельности. Он отражает основной принцип взаимосвязи организма и внешней среды, связывая их в единую систему, а представления о рефлекторной дуге объясняют механизм этой взаимосвязи. Основные положения рефлекторного принципа деятельности ЦНС были разработаны на протяжении примерно двух с половиной веков.
Согласно И.П. Павлову рефлекторная теория опирается на три основные принципа:
принцип детерминизма (причинности)- согласно которому рефлекторная реакция возникает только в ответ на раздражающий стимул. Принцип детерминизма устанавливает полную обусловленность материальными причинами всех явлений в организме, в том числе и высшей нервной деятельности. Изучение функций коры головного мозга позволило Павлову настолько точно познать законы, управляющие условно-рефлекторной деятельностью, что стало возможно в значительной степени управлять этой деятельностью у животных (собак) и заранее предсказывать, какие изменения произойдут при тех или иных условиях.
принцип структурности- устанавливает, что все нервные процессы являются результатом деятельности определенных структурных образований - нервных клеток, и зависят от свойств этих клеток. Однако, если до Павлова свойства различных клеток и клеточных групп центральной нервной системы рассматривались как постоянные, то Иван Петрович в учении об условных рефлексах показал, что свойства этих клеток изменяются в процессе развития. Локализация функций в коре головного мозга не должна, поэтому трактоваться только как распределение в пространстве клеток с различными свойствами. Кроме того, он предусматривает, что рефлекторная реакция возможна только в том случае, если все компоненты рефлекторной дуги находятся в анатомически и физиологически целостном состоянии. В такой формулировке он известен как принцип целостности.
наконец принцип анализа и синтезаустанавливает, что каждая ответная реакция всегда адекватна качествам и природе воздействующего раздражителя. Согласно этого принципа в процессе рефлекторной деятельности происходит, с одной стороны, дробление окружающей природы на огромную массу отдельно воспринимаемых явлений, а с другой - превращение одновременно или последовательно действующих раздражителей (различного характера) в комплексные. Грубый анализ может быть осуществлен уже низшими отделами нервной системы, так как раздражения разных рецепторов, каждая группа которых воспринимает определенные воздействия среды, вызывает лишь определенные безусловные рефлексы. Однако высший анализ, благодаря которому возможно существование животного организма в постоянно изменяющейся среде, осуществляется корой головного мозга и основан на способности образовывать условные рефлексы, а также, на способности к дифференцировке раздражителей.
Рефлекс- ответная реакция организма на изменения внешней или внутренней среды, осуществляемая при обязательном участии центральной нервной системы, в ответ на раздражение рецепторов. За счет рефлексов происходит возникновение, изменение или прекращение какой-либо деятельности организма.
Нервный путь, по которому распространяется возбуждение при осуществлении рефлексов, называют рефлекторной дугой. Следует отметить, что рефлекторная дуга является функциональным, а не структурным образованием и не отражаетстроение нервной системы, а объясняет принцип ее работы. Поэтому в последнее время используется терминконцептуальная рефлекторная дуга.
Существуют простые и сложные рефлекторные дуги. Простая рефлекторная дуга образована всего двумя нейронами: первый — чувствительный и второй — двигательный. В сложных рефлекторных дугах между этими нейронами включены еще вставочные нейроны. (Рис. 1).
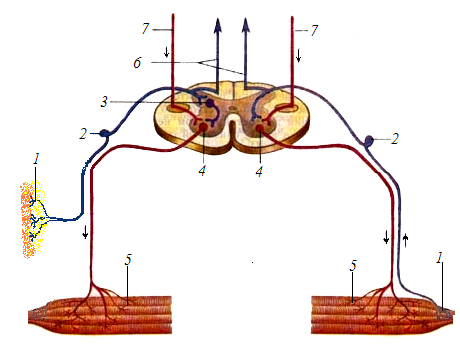
Рис. 1. Распространение нерв-ных импульсов по рефлекторной дуге (А – полисинаптической, Б – моносинаптической).
1 – рецептор;
2 – афферентный
(чувствительный) нейрон;
3 – вставочный нейрон;
4 – эфферентный
(двигательный) нейрон;
5 – эффектор;
6 – нервные волокна тонкого и
клиновидного пучков;
7 – волокна корково-спинно-
мозгового пути.
А Б
Классическая рефлекторная дуга состоит из пяти компонентов: 1) рецептора; 2) афферентного нервного пути; 3) рефлекторного центра; 4) эфферентного нервного пути; 5) эффектора (рабочего органа)
Рецептор- это чувствительное нервное окончание, воспринимающее раздражение, в котором энергия раздражителя превращается в энергию нервного импульса.
Рецептор может быть образован нервным окончанием чувствительного нейрона, который сам генерирует нервный импульс (первичночувствующие рецепторы) или представлен самостоятельной клеткой, которая деполяризуется с выделением медиатора действующего на синапсы чувствительного нейрона (вторичночувствующие рецепторы).
Различают: 1) экстерорецепторы - возбуждаются под влиянием раздражений из окружающей среды (рецепторы кожи, глаза, внутреннего уха, слизистой оболочки носа и ротовой полости); 2) интерорецепторы - воспринимают раздражения из внутренней среды организма (рецепторы внутренних органов, сосудов); 3) проприорецепторы - реагируют на изменение положения отдельных частей тела в пространстве (рецепторы мышц, сухожилий, связок, суставных сумок).
Определенные рефлекторные реакции могут включать различное количество рецепторов, афферентных и эфферентных нейронов и сложные процессы взаимодействия возбудительных и тормозных процессов в центрах нервной системы. Области тела, в пределах которых расположены специализированные рецепторы, раздражение которых обусловливает строго определенные рефлексы, получили название рефлексогенных зон или рецептивных полей.
Афферентный нервный путьпредставлен отростками рецепторных нейронов, несущих возбуждения в центральную нервную систему.
Афферентные нейроны(воспринимающие, чувствительные или центростремительные) передают информацию от рецепторов в центральную нервную систему. Тела этих нейронов расположены вне центральной нервной системы - в спинномозговых ганглиях и в ганглиях черепно-мозговых нервов.
Афферентный нейрон имеет ложноуниполярную форму, т. е. оба его отростка выходят из одного полюса клетки. Далее нейрон разделяется на длинный дендрит, образующий на периферии воспринимающее образование - рецептор, и аксон, входящий через задние рога в спинной мозг. К афферентным нейронам относят также нервные клетки, аксоны которых составляют восходящие пути спинного и головного мозга.
Рефлекторный центр (нервный центр)состоит из группы нейронов, расположенных на различных уровнях центральной нервной системы и передающих нервные импульсы с афферентного на эфферентный нервный путь. В качестве элементарного рефлекторного центра может рассматриваться вставочный нейрон.
Вставочные нейроны(интернейроны, контактные или промежуточные) - это, как правило, более мелкие клетки, осуществляющие связь между различными (в частности, афферентными и эфферентными) нейронами. Они передают нервные влияния в горизонтальном направлении (например, в пределах одного сегмента спинного мозга) и в вертикальном (например, из одного сегмента спинного мозга в другие - выше или нижележащие сегменты). Благодаря многочисленным разветвлениям аксона промежуточные нейроны могут одновременно возбуждать большое число других нейронов (например, звездчатые клетки коры).
Напомним, возможен вариант рефлекторной дуги и без вставочного нейрона, когда возбуждение с чувствительного нейрона передается непосредственно на эффекторный. Такая рефлекторная дуга называется двухнейронной или моносинаптической (Рис. 1 Б). Свойства таких рефлексов рассматриваются ниже.
Кроме того, описаны и местные рефлексы, замыкающиеся в периферических вегетативных ганглиях, т.е. не имеющие центрально звена. Именно они во многом восстанавливают и обеспечивают функции децентрализованных или пересаженных внутренних органов.
Эфферентный нервный путьпроводит нервные импульсы от центральной нервной системы к эффектору.
Эфферентные нейроны(исполнительные или центробежные) связаны с передачей нисходящих влияний от вышележащих этажей нервной системы к нижележащим или из центральной нервной системы к рабочим органам. Отростки этих нейронов образуют нисходящие пути спинного мозга.
Конечным путем эфферентных влияний являются расположенные в передних рогах спинного мозга тела двигательных нейронов или мотонейронов, от которых идут волокна к скелетным мышцам.
В боковых рогах спинного мозга находятся клетки вегетативной нервной системы, от которых идут пути к внутренним органам. Для эфферентных нейронов характерны разветвленная сеть дендритов и один длинный отросток - аксон.
Эффекторисполнительный орган, деятельность которого изменяется под влиянием нервных импульсов, поступающих к нему по образованиям рефлекторной дуги. Эффекторами могут быть мышцы или железы.
В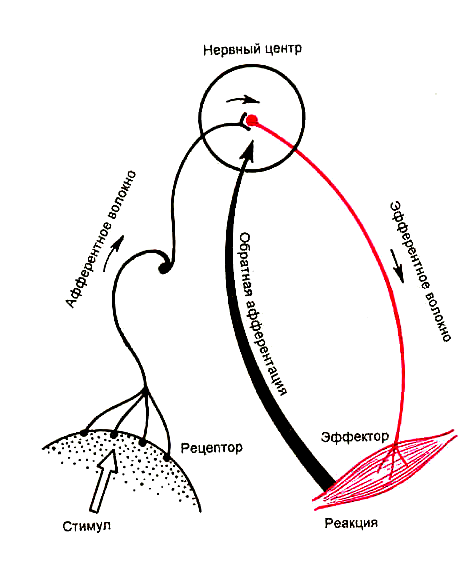 основе современного представления о
рефлекторной деятельности лежит понятиеполезного приспособительного
результата, ради которого совершается
любой рефлекс. Информация о достижении
полезного приспособительного результата
поступает в ЦНС по звену обратной связи
в видеобратной афферентации,которая
является обязательным компонентом
рефлекторной деятельности. В самом
простом случае это возбуждение которое
поступает от проприоцепторов сокращающейся
мышцы к вставочному нейрону этой
рефлекторной дуги и представляет собой
информацию о выполненном действии.
основе современного представления о
рефлекторной деятельности лежит понятиеполезного приспособительного
результата, ради которого совершается
любой рефлекс. Информация о достижении
полезного приспособительного результата
поступает в ЦНС по звену обратной связи
в видеобратной афферентации,которая
является обязательным компонентом
рефлекторной деятельности. В самом
простом случае это возбуждение которое
поступает от проприоцепторов сокращающейся
мышцы к вставочному нейрону этой
рефлекторной дуги и представляет собой
информацию о выполненном действии.
Принцип обратной афферентации введен в рефлекторную теорию П. К. Анохиным. Таким образом, по современным представлениям структурной основой рефлекса является не рефлекторная дуга, а рефлекторное кольцо,и к классической рефлекторной дуге добавляется еще один элемент (Рис.2) :
. Рис.2. Рефлекторное «кольцо»
В организации рефлекторного ответа важную роль может играть гуморальный механизм. Поступая из желез, являющихся эффекторами, в кровь, физиологически активные вещества (прежде всею медиаторы и гормоны) составляют в определенных условиях гуморальное звено рефлекторной дуги. Они передают в спинной и головной мозг соответствующую информацию, под влиянием которой возникает поток нервных импульсов из ЦНС в эффекторные органы. Классическая рефлекторная дуга, таким образом, усложняется, превращаясь в многозвенное кольцо, в котором нервное звено сменяется гуморальным, а гуморальное нервным. Принимая во внимание важное значение гистогематических, а также и гематоэнцефалических барьеров, можно говоритьо едином рефлекторном нейрогуморально - гормонально - барьерном механизме регуляции в организме
Классификация рефлексов. Уровни организации рефлекторных реакций.
Рефлексы отличаются большим многообразием.
Как уже отмечалось, в зависимости от числа синапсов в центральной части рефлекторной дуги (рефлекторном центре), различаютмоно- и полисинаптические рефлексы.
Наиболее простые - моносинаптическиерефлексы, обеспечиваемые двухнейронной рефлекторной дугой, образованной чувствительным и двигательным нейронами (Рис.1Б). В организме человека таких рефлексов немного и большинство из них - эторефлексы растяжения, вызываемые растяжением мышцы.
Рецепторная и эффекторная части рефлекторных дуг этих рефлексов находятся в мышце, поэтому этот рефлекс называют собственным рефлексом мышцы (миотатическим рефлексом). Рецепторы растяжения, расположены на так называемых внутриверетенных (интрафузальных)мышечных волокнах, на их средних, лишенных сократимости, но легко растяжимых частях. Эти же интрафузальные мышечные волокна на своих концах имеют моторную иннервацию из малых гамма - мотонейронов. Импульсы гамма-мотонейронов могут возбуждать и таким образом приводить в сокращенное состояние полярные зоны интрафузальных мышечных волокон, срединная (рецепторная) часть волокна при этом растягивается, чтов свою очередь вызывает моносинаптический рефлекс растяжения. Таким образом, порог возникновения коленного рефлекса и его сила могут регулироваться влияниями из ЦНС через гамма-мотонейроны.
Наиболее известным примером моносинаптического рефлекса растяжения является пателлярный (коленный) рефлекс, когда четырехглавую мышцу подвергают короткому растяжению в результате легкого постукивания по сухожилию ниже коленной чашечки. Следует иметь в виду, что выражение «сухожильный рефлекс» вводит в заблуждение, так как и в коленном и в других «сухожильных» рефлексах, реализуется моносинаптический рефлекс растяжения мышцы.
Отметим клинически важные примеры таких рефлекторных тестов: растяжение (путем постукивания по подбородку) мышцы, которая вызывает закрывание рта (рефлекс закрывания рта); растяжение двуглавой мышцы путем постукивания по ее сухожилию около локтя или удара по лучевой кости; растяжение трехглавой мышцы плеча путем постукивания по ее сухожилию чуть выше локтевого отростка; растяжение трехглавой мышцы голени путем постукивания по пяточному сухожилию (рефлекс ахиллова сухожилия) и др.
Моносинаптические рефлексы растяжения, вызываемые постукиванием по сухожилию, получили в клинике название Т-рефлексов.
Последовательное тестирование рефлексов растяжения имеет особое значение в связи с тем, что, их рефлекторные дуги проходят через разные сегменты спинного мозга, и нарушения определенных рефлексов могут показать, на каком уровне спинного мозга локализуется патологический процесс. Причем, клиническим показателем является не столько сила рефлекса, сколько наличие разницы между рефлексами с правой и с левой сторон, или аномалии некоторых рефлексов, выявляемые при их сравнении с общим рефлекторным поведением. Таким образом, регистрация Т-рефлексов служит простым тестом сохранности двигательных рефлексов, который в случае необходимости может дополняться другими диагностическими исследованиями.
Физиологическое значение таких рефлексов состоит в рефлекторном поддержании мышечной длины, что особенно важно для сохранения тонуса позных мышц. Наиболее наглядно это можно видеть на примере коленного рефлекса, где под тяжестью тела, стремящейся согнуть колено, сокращение мышцы как бы разгружает рецептор растяжения, что снижает рефлекс. Но даже небольшое ослабление мышцы сразу же усиливает нагрузку на рецептор и возвращает необходимую силу рефлекса. Подобные отношения в естественных условиях довольно часты.
Интересно, что если коленный и другие Т-рефлексы нижних конечностей оказываются ослабленными, то их можно усилить, попросив человека сцепить пальцы рук перед грудью в замок и пытаться тянуть руки в разные стороны или нажимать на руку другого человека (прием Ендрассика). Развиваемое при этом усилие вызывает облегченную активацию мотонейронов спинного мозга. Так маленькие дети, учась ходить, повышают устойчивость, очень крепко держась (благодаря врожденному хватательному рефлексу) за руку или палец мамы. Ослабление хватки приводит к ослаблению рефлекса, колени подгибаются, и ребенок практически вертикально опускается на ягодицы.
"Укороченным" вариантом коленного рефлекса (и других рефлексов растяжения) является так называемый Н-рефлекс мышцы (аш-рефлекс), возникающий при пороговом одиночном электрическом раздражении ее проприоцептивных афферентных волокон, при этом исключается собственно рецепторное звено. Изучение таких рефлексов имеет значение только в лабораторной и неврологической практике.
Все рефлексы, у которых число синапсов в ЦНС больше единицы, называют полисинаптическими. Это все остальные рефлексы (кроме моносинаптических),в их осуществлении участвует несколько (больше двух) последовательно включенных нейронов ЦНС.
Полисинаптические рефлексы очень многообразны, но классификация их на группы и типы, которая необходима здесь с дидактической точки зрения, не должна мешать пониманию того факта, что существует множество промежуточных форм, и что каждая признанная категория рефлексов является в том или ином отношении условной. Другой аспект, который легко упустить из виду, рассматривая отдельные рефлексы, независимо друг от друга, состоит в том, чтобольшинство мотонейронов и вставочных нейронов входит в очень многие рефлекторные дуги. Так, двигательный аксон мышц глотки участвует в рефлексах глотания, кашля, сосания, чихания и дыхания, т.е. образуетобщий конечный путьдля многочисленных рефлекторных дуг.
Существуют различные классификации рефлексов: по способам их вызывания, особенностям рецепторов, центральным нервным структурам их обеспечения, биологическому значению, сложности нейронной структуры рефлекторной дуги и т. д. Наиболее просто полисинаптические рефлексы классифицируются по элементам рефлекторной дуги.
По рецепторам, раздражение которых вызывает ответную реакцию, различаютэкстероцептивныерефлексы — рефлекторные реакции, инициируемые раздражением многочисленных экстерорецепторов (болевые, температурные, тактильные и т. д.),интероцептивныерефлексы (рефлекторные реакции, запускаемые раздражением интероцепторов: хемо-, баро-, осморецепторов и т.д.),проприоцептивныерефлексы (рефлекторные реакции, осуществляемые в ответ на раздражение проприорецепторов мышц, сухожилий, суставных поверхностей и т. д.).
По эффекторамразличают рефлексыдвигательные(реализуемые мышцами скелета),сердечные(проявляющиеся в изменениях работы мышцы сердца),сосудистые(проявляющиеся в изменении тонуса гладких мышц кровеносных сосудов),секреторные(реализуемые в развитии или изменении секреции желез) и т. п.
По локализации и характеру центрального звена различают рефлексы спинного мозга (спиномозговые) и рефлексы головного мозга (черепномозговые), среди которых, в свою очередь, можно выделить рефлексы продолговатого мозга, среднего мозга и др. (бульбарные, мезенцефальные, диэнцефальные, кортикальные рефлекторные реакции).
В зависимости от отдела нервной системы реализующей ответ то есть, отдела в котором расположен эфферентный нейрон, выделяют соматические и вегетативные рефлексы. Соматическая нервная система регулирует перемещение в пространстве (локомоцию), её эффекторами всегда являются скелетные мышцы. Вегетативная нервная система регулирует процессы жизнедеятельности, её эффекторы – гладкая мускулатура внутренних органов и железы.
По локализации рефлекторной дуги различаютцентральныерефлексы (дуга которых проходит через ЦНС) ипериферическиерефлексы, дуга которых замыкается вне ЦНС. Последними, могут быть только вегетативныерефлексы, они подразделяются на интраорганные и экстраорганные (в последнем случае рефлекс осуществляется с участием нейроновэкстраорганных ганглиев). Выделяют также межорганные рефлексы, которые могут быть и периферическими и центральными. Например, механическое раздражение желудкаможет затормозить сердечную деятельность.
Особым случаем периферических рефлексов являются, так называемые аксон - рефлексы. Аксон – рефлекс – общее название рефлексов осуществляющихся по разветвлениям аксона без участия тела нейрона. Рефлекторная дуга аксон – рефлекса не содержит синапсов и тел нейронов, возбуждение идет афферентно по одной ветви аксона, затем передается на другую ветвь и эфферентно распространяется к эффекторному органу. С помощью аксон – рефлексов может происходить регуляция внутренних органов и сосудов в определенной степени независимо от ЦНС,
По биологическому значению рефлексыделят на пищедобывательные, половые, защитные (оборонительные), исследовательские, Примером исследовательского рефлекса является ориентировочный рефлекс (рефлекс «что такое?») — поворот головы, глаз, туловища в сторону внезапно появившегося раздражителя. Ориентировочные рефлексы лежат в основе процесса научения.
Изучение роли нервной системы в организации поведения, привело к еще одному подходу в классификации рефлексов.
Все рассмотренные выше рефлексы наблюдаются примерно в одинаковой форме у всех особей данного вида и не требуют специальных условий для проявления. Такие стереотипные реакции в значительной степени предопределены структурной организацией нервной системы, и, прежде всего, организацией рефлекторных дуг, которые находятся в филогенетически древних отделах центральной нервной системы - спинном мозгу и стволе мозга, даже если рефлексы являются очень сложными. Такие рефлексы передаются по наследству и проявляются сразу после рождения или по мере созревания нервных структур, через которые они реализуются, поэтому они получили название врожденных. А поскольку для их проявления не требуется специальных условий (кроме действия раздражителя), то их называютбезусловными.
Однако постепенно стало ясно, что организм с высокоорганизованной нервной системой, способен обучаться рефлекторным реакциям, что помогает ему лучше и с меньшими усилиями отвечать на постоянно меняющиеся условия внешней среды. Такие рефлексы назвали приобретенными. Рефлекторные дуги таких приобретенных рефлексов (которые могут снова забываться) обычно находятся на высших (корковых) уровнях центральной нервной системы. Хорошо знакомыми примерами приобретенных рефлексов, которые подвергались тщательному экспериментальному изучению, являютсяусловные рефлексы, а также изменения поведения, вызываемыеоперантными условными рефлексами.
Механизмы возникновения уловных рефлексов и их роль в организации поведения является предметом изучения физиологии высшей нервной деятельности.
С учетом уровня эволюционного развития, совершенствования сложности нервного субстрата, обеспечивающего соответствующую рефлекторную реакцию, физиологического значения, уровня интегративной деятельности организм а выделяют шесть основных видов рефлексов, или уровней рефлекторных реакций (А. Б. Коган):
Элементарные безусловные рефлексыпредставлены простыми рефлекторными реакциями, осуществляемыми на уровне отдельных сегментов спинного мозга. Они имеют местное значение, вызываются локальным раздражением рецепторов данного сегмента тела и проявляются в виде локальных сегментарных сокращений поперечнополосатой мускулатуры. Элементарные безусловные рефлексы осуществляются по жестко детерминированным программам и имеют четкую определенную структурную основу в виде сегментарного аппарата спинного мозга. В результате такие рефлекторные реакции отличаются высокой степенью автоматизма и стереотипности. Функциональная роль этой категории рефлексов заключается в обеспечении простейших приспособительных реакций к внешним воздействиям местного значения, а также в приспособительных изменениях отдельных внутренних органов.
Координационные безусловные рефлексыпредставляют собой согласованные акты локомоторной деятельности или комплексные реакции вегетативных функциональных объединений внутренних органов. Эти рефлексы также вызываются раздражением определенных групп внешних или внутренних рецепторов, однако их эффект не ограничивается локальной реакцией путем последующей активации широкого класса экстеро-, ицтеро- и проприорецепторов, а формирует сложные координационные акты сокращения и расслабления, возбуждения или торможения деятельности ряда внутренних органов.
В физиологических механизмах реализации рефлекторных реакций этого типа значительное место занимает принцип обратной связи, обеспечиваемый соответствующими спинномозговыми структурами и осуществляющий антагонистическую, реципрокную иннервацию мышц-синергистов и антагонистов. Функциональное назначение координационных безусловных рефлексов — формирование на базе локальных элементарных безусловных рефлексов целостных, целенаправленных локомоторных актов или гомеостатических систем организма.
Интегративные безусловные рефлексыпредставляют собой дальнейший шаг в интеграции отдельных безусловных рефлексов, осуществляющих сложные двигательные локомоторные акты организма в тесной связи с вегетативным обеспечением, формируя тем самым комплексные поведенческие акты, имеющие определенное биологическое значение. Рефлекторные реакции этого типа инициируются такими биологически важными стимулами, как пищевые, болевые раздражители. Определяющим на входе этих рефлекторных актов становятся не физико-химические свойства стимулов, а в первую очередь их биологическое значение. Интегративные безусловные рефлексы всегда носят целостный системный характер, включая достаточно выраженные соматические и вегетативные компоненты. Их реализация оказывается весьма пластичной, тесно связанной со многими сильно развитыми проприоцептивными обратными связями, обеспечивающими точную коррекцию выполняемого сложного поведенческого акта в соответствии с изменениями в состоянии организма. Пример такой реакции — ориентировочная реакция. Биологическое значение последней заключается в перестройке организма, которая обеспечивает оптимальную подготовку к восприятию и быстрому анализу нового неизвестного сигнала в целях организации рационального ответа. Интегративные безусловные рефлексы требуют для своей реализации надсегментарных механизмов нервно-рефлекторной регуляции поведения организма. Эти рефлексы означают переход от сравнительно простых безусловных рефлексов к поведенческим актам.
Сложнейшие безусловные рефлексы (инстинкты)представляют собой видовые стереотипы поведения, организующиеся на базе интегративных рефлексов по генетически заданной программе. В качестве запускающих стереотипные поведенческие реакции раздражений выступают стимулы, имеющие отношение к питанию, защите, размножению и другим биологически важным потребностям организма.
Сложнейшие безусловные рефлексы образованы последовательными интегративными реакциями, построенными таким образом, что завершение одной реакции становится началом следующей. Адаптивность инстинктов усиливается благодаря наслоению на сложнейшие безусловные рефлексы условных, приобретаемых на ранних этапах онтогенеза. Нервный субстрат, ответственный за физиологические механизмы инстинктивного поведения, представляет иерархическую систему соподчиненных центров интегративных, координационных и элементарных безусловных рефлексов. Жесткая предопределенность инстинктивных реакций обусловлена этапной последовательностью актов инстинктивного поведения, ограничивающей сферу функционирования обратной связи от последующего этапа к предыдущему, уже реализованному. Инстинктивные реакции отражают исторический опыт вида. В субъективной сфере человека сложнейшие безусловные рефлексы проявляются в виде последовательных влечений и желаний, в сложной игре эмоций.
Элементарные условные рефлексыпроявляются в интегративных реакциях, вызываемых ранее индифферентными раздражителями, приобретающими сигнальное значение в результате жизненного опыта или подкрепления их безусловными стимулами (сигналами), имеющими биологическое значение. Основным принципиальным отличием этой категории рефлекторных реакций является то, что они образуются в процессе индивидуальной жизни. Условно-рефлекторные реакции образуются, усложняются, видоизменяются на протяжении всей жизни; наиболее простые из них формируются в раннем возрасте. Нервным субстратом, отвечающим за осуществление условно-рефлекторных реакций, является филогенетически наиболее молодая структура головного мозга — кора больших полушарий. Многоканальность и взаимозаменяемость путей реализации условно-рефлекторного механизма лежат в основе высокой пластичности и надежности условно-рефлекторных реакций. В системе рефлекторных реакций появление условного рефлекса означает качественно новый скачок в приспособительном поведении высших животных и человека. Условно-рефлекторные реакции дают возможность организму заблаговременно отвечать на приближающиеся жизненно важные ситуации. В психической сфере деятельности человека условные рефлексы закладывают начало ассоциативному способу мышления.
Сложные формы высшей нервной деятельностипредставлены психическими реакциями, возникающими на основе интеграции элементарных условных рефлексов и аналитико-синтетических механизмов абстрагирования. Абстрагирование от конкретного содержания безусловных подкрепляющих раздражителей обеспечивает возможность более полного и целостного восприятия окружающего мира, адекватного прогнозирования и программирования поведения. В качестве вызывающих подобные реакции стимулов обычно выступают сложные комплексные раздражители. Часто такие рефлекторные реакции имеют усеченную рефлекторную дугу (отсутствует эфферентное звено рефлекторной дуги). Сложные формы высшей нервной деятельности оказываются связанными с синтетическими процессами, обеспечивающими целостные субъективные образы внешнего мира, целенаправленные программы поведения, различные проявления абстрактной мыслительной деятельности человека (психонервная деятельность, рассудочное мышление, функции второй сигнальной системы).
Нервные центры. Свойства нервных центров.
От рецепторов нервные импульсы по афферентным путям поступают в нервные центры. Следует различать анатомическое и физиологическое понимание нервного центра.
Анатомическое определениенервного центра. Нервный центр это совокупность нейронов, расположенных в определенном отделе центральной нервной системы. За счет работы такого нервного центра осуществляется несложная рефлекторная деятельность, например коленный рефлекс. Нервный центр этого рефлекса располагается в поясничном отделе спинного мозга.
Физиологическое определениенервного центра. Нервный центр это сложное функциональное объединение нескольких анатомических нервных центров, расположенных на разных уровнях центральной нервной системы и обусловливающих за счет своей активности сложнейшие рефлекторные акты. Например, в осуществлении пищевых реакций участвуют многие органы (железы, мышцы, кровеносные и лимфатические сосуды и т. л.). Деятельность этих органов регулируется нервными импульсами, поступающими из нервных центров, располагающихся в различных отделах центральной нервной системы. При пищевых реакциях различные анатомические нервные центры функционально объединяются для получения определенного полезного результата.
Физиологические свойства нервных центров. Нервные центры обладают рядом характерных функциональных свойств, зависящих от наличия синапсов и большого количества нейронов, входящих в их состав. Основными свойствами нервных центров являются:
Одностороннее проведение возбуждения.В центральной нервной системе возбуждение распространяется только в одном направлении от рецепторного нейрона к эффекторному. Это обусловлено наличием в нервных центрах синапсов, в которых передача возбуждения возможна только в одном направлении от нервного окончания, выделяющего медиатор, к постсинаптической мембране.
Задержка проведения возбуждения в нервных центрахтакже связана с наличием большого количества синапсов. На выделение медиатора, его диффузию через синаптическую щель, возбуждение постсинаптической мембраны требуется больше времени, чем на распространение возбуждения по нервному волокну.
Суммация возбуждений в нервных центрахвозникает или при нанесении слабых, но повторяющихся (ритмичных) раздражений, или при одновременном действии нескольких подпороговых раздражений. Механизм этого явления связан с накоплением медиатора на постсинаптической мембране и повышением возбудимости клеток нервного центра. Примером суммации возбуждения может служить рефлекс чихания. Этот рефлекс возникает только при длительном раздражении рецепторов слизистой оболочки носа.
Трансформация ритма возбуждений. Центральная нервная система на любой ритм раздражения, даже медленный, отвечает залпом импульсов. Частота возбуждений, поступающих из нервных центров на периферию к рабочему органу, колеблется от 50 до 200 в 1 с. Этой особенностью центральной нервной системы объясняется то, что все сокращения скелетных мышц в организме являются тетаническими.
Рефлекторное последействие. Рефлекторные акты заканчиваются не одновременно с прекращением вызвавшего их раздражения, а через некоторый, иногда сравнительно длительный период. Это явление получило название рефлекторного последействия. Установлены два механизма, обусловливающие последействие. Первый связан с тем, что возбуждение в нервных клетках исчезает не сразу после прекращения раздражения. В течение некоторого времени (сотые доли секунды) нервные клетки продолжают давать ритмические разряды импульсов. Этот механизм может обусловить лишь сравнительно кратковременное последействие. Второй механизм является результатом циркуляции нервных импульсов по замкнутым нейронным цепям нервного центра и обеспечивает более длительное последействие.
Возбуждение одного из нейронов передается на другой, а по ответвлениям его аксона вновь возвращается к первой нервной клетке и т. д. Циркуляция нервных импульсов в нервном центре будет продолжаться до тех пор, пока не наступит утомление одного из синапсов или же активность нейронов не будет приостановлена приходом тормозных импульсов.
Утомление нервных центров. Нервные центры в отличие от нервных волокон легко утомляемы. При продолжительном раздражении афферентных нервных волокон утомление нервного центра проявляется постепенным снижением, а затем и полным прекращением рефлекторного ответа.
Эта особенность нервных центров доказывается следующим образом. После прекращения мышечного сокращения в ответ на раздражение афферентных нервов начинают раздражать эфферентные волокна, иннервирующие мышцу. В этом случае мышца вновь сокращается. Следовательно, утомление развилось не в эфферентных путях, а в нервном центре.
В многочисленных исследованиях установлено, что наиболее утомляемыми являются воспринимающие нейроны (чувствительные и промежуточные) по сравнению с эфферентными нервными клетками рефлекторной дуги. В настоящее время считают, что утомление нервных центров связано прежде всего с нарушением передачи возбуждения в синапсах. Такое нарушение может быть обусловлено уменьшением запасов медиатора или снижением чувствительности к медиатору постсинаптической мембраны нервной клетки.
Принципы координации в деятельности ЦНС.
В условиях физиологической нормы работа всех органов и систем тела является согласованной: на воздействия из внешней и внутренней среды организм реагирует как единое целое. Согласованное проявление отдельных рефлексов, обеспечивающих выполнение целостных рабочих актов, носит название координации.
Явления координации играют важную роль в деятельности двигательного аппарата. Координация таких двигательных актов, как ходьба или бег, обеспечивается взаимосвязанной работой нервных центров.
За счет координированной работы нервных центров осуществляется совершенное приспособление организма к условиям существования. Это происходит не только за счет деятельности двигательного аппарата, но и за счет изменений вегетативных функций организма (процессов дыхания, кровообращения, пищеварения, обмена веществ и т.д.).
Установлен ряд общих закономерностей - принципов координации: принцип конвергенции; принцип иррадиации возбуждения; принцип реципрокности; принцип последовательной смены возбуждения торможением и торможения возбуждением; феномен «отдачи»; цепные и ритмические рефлексы; принцип общего конечного пути; принцип обратной связи; принцип доминанты.
Разберем некоторые из них.
Принцип конвергенции. Этот принцип установлен английским физиологом Шеррингтоном. Импульсы, при ходящие в центральную нервную систему по различным афферентным волокнам, могут сходиться (конвергировать) к одним и тем же вставочным и эфферентным нейронам. Конвергенция нервных импульсов объясняется тем, что афферентных нейронов в несколько раз больше, чем эфферентных, поэтому афферентные нейроны образуют на телах и дендритах эфферентных и вставочных нейронов многочисленные синапсы.
Принцип иррадиации. Импульсы, поступающие в центральную нервную систему при сильном и длительном раздражении рецепторов, вызывают возбуждение не только данного рефлекторного центра, но и других нервных центров. Это распространение возбуждения в центральной нервной системе получило название иррадиации. Процесс иррадиации связан с наличием в центральной нервной системе многочисленных ветвлений аксонов и особенно дендритов нервных клеток и цепей вставочных нейронов, которые объединяют друг с другом различные нервные центры.
Принцип реципрокности(сопряженности) в работе нервных центров. Суть его заключается в том, что при возбуждении одних нервных центров деятельность других может затормаживаться. Принцип реципрокности был показан по отношению к нервным центрам мышц-антагонистов - сгибателей и разгибателей конечностей. Только при такой взаимосочетанной (реципрокной) иннервации возможен акт ходьбы.
Наиболее отчетливо он проявляется у животных с удаленным головным мозгом и сохраненным спинным (спинальное животное), но может происходить сопряженное, реципрокное торможение и других рефлексов. Под влиянием головного мозга реципрокные отношения могут изменяться. Человек или животное в случае необходимости может сгибать обе конечности, совершать прыжки и т. д.
Реципрокные взаимоотношения центров головного мозга определяют возможность человека овладеть сложными трудовыми процессами и не менее сложными специальными движениями, совершающимися при плавании, акробатических упражнениях и пр.
Принцип общего конечного пути. Этот принцип связан с особенностью строения центральной нервной системы. Эта особенность, как уже указывалось, состоит в том, что афферентных нейронов в несколько раз больше, чем эфферентных, в результате чего различные афферентные импульсы сходятся к общим выходящим путям.
Количественные соотношения между нейронами схематически можно представить в виде воронки: возбуждение вливается в центральную нервную систему через широкий раструб (афферентные нейроны) и вытекает из нее через узкую трубку (эфферентные нейроны). Общими путями могут быть не только конечные эфферентные нейроны, но и вставочные.
Импульсы, сходящиеся в общем пути, «конкурируют» друг с другом за использование этого пути. Так достигается упорядочение рефлекторного ответа, соподчинение рефлексов и затормаживание менее существенных. Вместе с тем организм получает возможность реагировать на различные раздражения из внешней и внутренней среды при помощи сравнительно небольшого количества исполнительных органов.
Принцип обратной связи. Этот принцип изучен И. М. Сеченовым, Шеррингтоном, П. К. Анохиным и рядом других исследователей. При рефлекторном сокращении скелетных мышц возбуждаются проприорецепторы. От проприорецепторов нервные импульсы несущие информацию о характеристиках этого мышечного сокращения вновь поступают в центральную нервную систему. Этим контролируется точность совершаемых движений. Подобные афферентные импульсы, возникающие в организме в результате рефлекторной деятельности органов и тканей (эффекторов), получили названиевторичных афферентных импульсов, илиобратной связи.
Обратные связи могут быть положительными и отрицательными. Положительные обратные связи способствуют усилению рефлекторных реакций, отрицательные - их угнетению. За счет положительных и отрицательных обратных связей осуществляется, например, регуляция относительного постоянства величины артериального давления.
Принцип доминанты.Принцип доминанты сформулирован А. Л. Ухтомским, Этот принцип играет важную роль в согласованной работе нервных центров.Доминанта - это временно господствующий очаг возбуждения в центральной нервной системе, определяющий характер ответной реакции организма на внешние и внутренние раздражения.
Доминантный очаг возбуждения характеризуется следующими основными свойствами:
повышенной возбудимостью;
стойкостью возбуждения;
способностью к суммированию возбуждения;
инерцией, доминанта в виде следов возбуждения может длительно сохраняться и после прекращения вызвавшего ее раздражения.
Доминантный очаг возбуждения способен притягивать (привлекать) к себе нервные импульсы из других нервных центров, менее возбужденных в данный момент. За счет этих импульсов активность доминанты еще больше увеличивается, а деятельность других нервных центров подавляется.
Доминанты могут быть экзогенного и эндогенного происхождения. Экзогенная доминанта возникает под влиянием факторов окружающей среды. Например, при чтении интересной книги человек может не слышать звучащую в это время по радио музыку.
Эндогенная доминанта возникает под влиянием факторов внутренней среды организма, главным образом гормонов и других физиологически активных веществ. Например, при понижении содержания питательных веществ в крови, особенно глюкозы, происходит возбуждение пищевого центра, что является одной из причин пищевой установки организма животных и человека.
Доминанта может быть инертной (стойкой), и для ее разрушения необходимо возникновение нового более мощного очага возбуждения.
Доминанта лежит в основе координационной деятельности организма, обеспечивая поведение человека и животных в окружающей среде, а также эмоциональных состояний, реакций внимания. Формирование условных рефлексов и их торможение также связано с наличием доминантного очага возбуждения.
Спинной мозг.
Спинной мозг лежит в позвоночном канале и представляет собой тяж длиной 41 - 45 см (у взрослого), несколько сплющенный спереди назад. Вверху он непосредственно переходит в головной мозг, а внизу заканчивается заострением - мозговым конусом - на уровне II поясничного позвонка.
Характерной чертой организации спинного мозга является периодичность его структуры в форме сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов (серое вещество) и выходы в виде задних корешков. Морфологической границы между сегментами спинного мозга не существует, поэтому деление на сегменты является функциональным. Каждый сегмент через свои корешки иннервирует три метамера тела собственный, половину вышележащего и половину нижележащего, а каждый метамер тела получает иннервацию от трех сегментов спинного мозга. Такое устройство гарантирует осуществление функций спинного мозга при возможных его перерывах и других поражениях.
Распределение функций входящих и выходящих волокон спинного мозга подчиняется определенному закону: все чувствительные (афферентные) волокна входят в спинной мозг через его задние корешки, а двигательные и вегетативные (эфферентные) выходят через передние корешки.Этот закон известен как закон Белла-Мажанди.
В задних корешках волокон гораздо больше, чем в передних (их соотношение у человека примерно 5:1) т. е. при большом разнообразии поступающей информации организм использует незначительное количество исполнительных приборов. По задним корешкам в спинной мозг поступают импульсы от рецепторов скелетных мышц, сухожилий, кожи, сосудов, внутренних органов. Передние корешки содержат волокна к скелетным мышцам и вегетативным ганглиям.
Задние корешкиобразованы волокнами одного из отростков афферентных нейронов, тела которых расположены вне центральной нервной системы – в межпозвоночных ганглиях, а волокна другого отростка связаны с рецептором. Общее число афферентных волокон у человека достигает примерно 1 млн. Они различаются по диаметру. Наиболее толстые идут от рецепторов мышц и сухожилий, средние по толщине—от тактильных рецепторов кожи, от части мышечных рецепторов и от рецепторов внутренних органов (мочевого пузыря, желудка, кишечника и др.), наиболее тонкие миелинизированные и немиелинизированные волокна—от болевых рецепторов и терморецепторов. Одна часть афферентных волокон заканчивается на нейронах спинного мозга, другая часть направляется к нейронам продолговатого мозга, образуя спинно-бульбарный путь.
Передние корешкисостоят из отростков мотонейронов передних рогов спинного мозга и нейронов боковых рогов. Волокна первых направляются к скелетной мускулатуре, а волокна вторых переключаются в вегетативных ганглиях на другие нейроны и иннервируют внутренние органы.
Спинной мозг имеет два утолщения: шейное и поясничное, соответствующие местам выхода из него нервов, идущих к верхней и нижней конечностям. Передней срединной щелью и задней срединной бороздкой спинной мозг делится на две симметричные половины, каждая в свою очередь имеет по две слабовыраженные продольные борозды, из которых выходят передние и задние корешки - спинномозговые нервы. Эти борозды разделяют каждую половину на три продольных тяжа - канатика: передний, боковой и задний. В поясничном отделе корешки идут параллельно концевой нити и образуют пучок, носящий название конского хвоста.
В нутри
спинной мозг состоит из серого и белого
вещества. Серое вещество заложено внутри
и со всех сторон окружено белым. В каждой
из половин спинного мозга оно образует
два неправильной формы вертикальных
тяжа с передними и задними выступами -
столбами, соединенных перемычкой -
центральным промежуточным веществом,
в середине которого заложен центральный
канал, проходящий вдоль спинного мозга
и содержащий спинномозговую жидкость.
В грудном и верхнем поясничном отделах
имеются также боковые выступы серого
вещества. Таким образом, в спинном мозге
различают три парных столба серого
вещества: передний, боковой и задний,
которые на поперечном разрезе спинного
мозга носят название переднего, бокового
и заднего рогов.
нутри
спинной мозг состоит из серого и белого
вещества. Серое вещество заложено внутри
и со всех сторон окружено белым. В каждой
из половин спинного мозга оно образует
два неправильной формы вертикальных
тяжа с передними и задними выступами -
столбами, соединенных перемычкой -
центральным промежуточным веществом,
в середине которого заложен центральный
канал, проходящий вдоль спинного мозга
и содержащий спинномозговую жидкость.
В грудном и верхнем поясничном отделах
имеются также боковые выступы серого
вещества. Таким образом, в спинном мозге
различают три парных столба серого
вещества: передний, боковой и задний,
которые на поперечном разрезе спинного
мозга носят название переднего, бокового
и заднего рогов.
Передний рогимеет округлую или четырехугольную форму исодержит клетки, дающие начало передним (двигательным) корешкам спинного мозга.
Задний рогуже и длиннее и включает клетки, к которым подходят чувствительные волокна задних корешков.
Боковой рог образует небольшой треугольной формы выступ, состоящий из клеток, относящихся к вегетативной части нервной системы.
Белое вещество спинного мозга составляет передний, боковой и задний канатики и образовано преимущественно продольно идущими нервными волокнами, объединенными в пучки - проводящие пути.
Среди них выделяют три основных вида:
волокна, соединяющие участки спинного мозга на различных уровнях;
двигательные (нисходящие) волокна, идущие из головного мозга в спинной на соединение с клетками, дающими начало передним двигательным корешкам;
чувствительные (восходящие) волокна, которые частично являются продолжением волокон задних корешков, частично отростками клеток спинного мозга и восходят кверху к головному мозгу.
От спинного мозга, образуясь из передних и задних корешков, отходит 31 пара смешанных спинномозговых нервов: 8 пар шейных, 12 пар грудных, 5 пар поясничных, 5 пар крестцовых и 1 пара копчиковых. Участок спинного мозга, соответствующий отхождению пары спинномозговых нерввов, называют сегментом спинного мозга. В спинном мозге выделяют 31 сегмент.
Спинной мозг выполняет две функции: рефлекторную и проводниковую.
Рефлекторные центры спинного мозга. Объем функций, осуществляемых спинным мозгом, чрезвычайно велик. В шейном отделе спинного мозга находятся центр диафрагмального нерва и центр сужения зрачка, в шейном и грудном отделах - центры мышц верхних конечностей, мышц груди, спины и живота, в поясничном отделе - центры мышц нижних конечностей, в крестцовом отделе - центры мочеиспускания, дефекации и половой деятельности, в боковых рогах грудного и поясничного отделов спинного мозга - центры потоотделения и спинальные сосудодвигательные центры.
В естественных условиях эти рефлексы всегда испытывают влияние высших отделов головного мозга. Степень проявления рефлексов зависит от того, сохраняются ли связи структур спинного мозга со структурами головного мозга. После децеребрации (удаления головного мозга) или спинализации (отделения спинного мозга от головного посредством перерезки) исчезают многие сложные формы активности, создаваемые спинным мозгом. При этом определенное значение принадлежит уровню организации подопытного животного. Например, спинальная лягушка, может сидеть и вырываться, когда ее схватывают, собака же сама не может ни стоять, ни ходить. Причина этого в разобщении спинного мозга и вышележащих структур нарушает. При этом, в частности, исчезают периодические разряды дыхательных мышц, обеспечивающие дыхательные движения, пропадают тонические разряды симпатических нейронов, поддерживающих сосудистый тонус и соответственно артериальное давление.
В зависимости от числа нейронов, участвующих в проведении возбуждения, рефлекторные дуги спинного мозга делятся на моносинаптические и полисинаптические. За исключением моносинаптических рефлексов растяжения рефлекторные дуги всех остальных спиномозговых рефлексов являются полисинаптическими.
К числу рефлексов спинного мозга относятся
защитные рефлексы, рефлексы на растяжение,
мышц-антагонистов, висцеромоторные,
вегетативные рефлексы. Эта классификация
весьма условна, она указывает только
на многообразие рефлекторных ответов.
Даже у спинального животного трудно
встретить рефлексы, которые относились
бы только к одной из названных групп.
числу рефлексов спинного мозга относятся
защитные рефлексы, рефлексы на растяжение,
мышц-антагонистов, висцеромоторные,
вегетативные рефлексы. Эта классификация
весьма условна, она указывает только
на многообразие рефлекторных ответов.
Даже у спинального животного трудно
встретить рефлексы, которые относились
бы только к одной из названных групп.
Защитные рефлексызапускаются, как правило, с рецепторов кожи, хотя характер ответной реакции сильно зависит от силы и вида раздражителя. Чаще всего конечная реакция выглядит в виде усиления сокращения мышц сгибателей.
Рефлексы растяженияпроявляются укорочением мышцы в ответ на ее растяжение.
Основными рецепторами в этом случае служат нервно-мышечные веретена, афферентным звеном чувствительные волокна соматических нервов и дорсальных корешков спинного мозга. Эти рефлекторные дуги чаще всего замыкаются в спинном мозге. Начало и конец рефлекторной дуги связаны с мышцей. Рефлексы наиболее выражены в мышцах-разгибателях. Для того. чтобы организм мог противостоять силе земного притяжения, эти мышцы должны находиться в состоянии тонического напряжения. физиологическое значение этих рефлексов состоит в том, что они участвуют в сохранении статики и положения тела, регулируя степень сокращения мышцы в соответствии с падающими на нее раздражениями.
Рефлексы мышц-антагонистовлежат в основе локомоторных актов и характеризуются тем, что при возбуждении мотонейронов сгибателей одновременно происходит торможение мотонейронов мыщц-разгибателей. При этом в конечности другой стороны наблюдаются обратные явления. В целом это создает правильное чередование противоположных по функциональному значению мышечных сокращений. Механизм, обусловливающий такое чередование активности различных двигательных ядер, например при ходьбе, локализуется в интернейронном аппарате спинного мозга. Вместе с тем для его активации необходимо поступление тонического нисходящего сигнала из двигательных центров головного мозга.
Висцеромоторные рефлексывозникают при возбуждении афферентных волокон внутренних органов и характеризуются появлением двигательных реакций мышц грудной и брюшной стенки, мышц-разгибателей спины. Возникновение этих рефлексов связано с существованием конвергенции висцеральных и соматических аффферентных волокон к одним и тем же интернейронам спинного мозга.
Вегетативные рефлексызаключаются, во-первых, в появлении полисинаптических разрядов в преганглионарных симпатических волокнах в ответ на возбуждение симпатических и соматических чувствительных клеток, во-вторых, в возникновении рефлекторных реакций парасимпатических нейронов в ответ на раздражение чувствительных путей.
Наряду с выполнением собственных рефлекторных реакций нейронные структуры спинного мозга служат аппаратом для реализации большого числа сложных процессов, осуществляемых различными отделами головного мозга.
Проводниковая функция спинного мозга. Через спинной мозг проходят восходящие и нисходящие нервные пути.
Восходящие нервные путипередают информацию от тактильных, болевых, температурных рецепторов кожи и от проприорецепторов мышц через нейроны спинного мозга в вышележащие отделы центральной нервной системы, к мозжечку и горе головного мозга.
Нисходящие нервные пути (пирамидный и экстрапирамидный) связывают кору головного мозга, подкорковые ядра и образования ствола мозга с мотонейронами спинного мозга. Они обеспечивают влияние высших отделов ЦНС на деятельность скелетных мышц.
Продолговатый мозг и мост
Непосредственным продолжением спинного мозга у всех позвоночных животных и человека является продолговатый мозг. Продолговатый мозг и варолиев мост (мост мозга) объединяют под общим названиемзаднего мозга.
Функции продолговатого мозга были изучены на бульбарных животных, у которых поперечным разрезом продолговатый мозг отделен от среднего. Следовательно, жизнь бульбарных животных осуществляется за счет деятельности спинного и продолговатого мозга. У таких животных отсутствуют произвольные движения, отмечается потеря всех видов чувствительности, нарушается регуляция температуры тела (теплокровное животное превращается в холоднокровное). У бульбарных животных сохраняются рефлекторные реакции организма и осуществляется регуляция функций внутренних органов.
В продолговатом мозге по сравнению со спинным мозгом нет четкого сегментарного распределения серого и белого вещества. Скопление нервных клеток приводит к образованию ядер, являющихся центрами более или менее сложных рефлексов. Из 12 пар черепных нервов, связывающих головной мозг с периферией организма - его рецепторами и эффекторами, восемь пар (V-ХII) берут свое начало в заднем мозге. В продолговатом мозгу и мосте расположены ядра следующих черепных нервов:
пара V черепных нервов – тройничный нерв, имеет двигательное и чувствительное ядра. Двигательное ядро расположено в мосту, иннервирует жевательные мышцы и вызывает движения нижней челюсти а также напрягает мягкое небо и барабанную перепонку. Чувствительные ядра (среднемозговое, мостовое, спинальное) получают от кожи, слизистых оболочек, органов лица и головы тактильную, температурную, висцеральную, проприоцептивную, болевую импульсацию, входят в проводниковый отдел соответствующих анализаторов и участвуют в различных рефлексах (например, жевательном, глотательном, чихательном).
пара VI черепных нервов – отводящий нерв; ядро отводящего нерва расположено в мосту. Иннервирует наружную прямую мышцу глаза, вызывает поворот его кнаружи.
пара VII черепных нервов – лицевой нерв; ядра лицевого нерва находятся в мосту. Двигательное ядро вызывает сокращения мимической и вспомогательной жевательной мускулатуры, регулирует передачу звуковых колебаний в среднем ухе в результате сокращения стременной мышцы. Чувствительное ядро одиночного пути, иннервируя вкусовые луковицы передних 2/3 языка, анализирует вкусовую чувствительность, участвует в моторных и секреторных пищеварительных рефлексах. Верхнее слюноотделительное (парасимпатическое) ядро стимулирует выделение секретов подъязычной, подчелюстной слюнных и слезной желез.
пара VIII черепных нервов — преддверно-улитковый нерв; его чувствительные ядра расположены в продолговатом мозге. Вестибулярные ядра, иннервируя рецепторы вестибулярного аппарата, участвуют в регуляции позы и равновесия тела (статические и статокинетические рефлексы), в вестибулоглазных и вестибуловегетативных рефлексах, входят в проводниковый отдел вестибулярного анализатора. Улитковые ядра, иннервирующие слуховые рецепторы, участвуют в слуховом ориентировочном рефлексе, входят в проводниковый отдел слухового анализатора.
пара IX — языкоглоточный нерв; ядра языкоглоточного нерва расположены в продолговатом мозге. Двойное (двигательное) ядро вызывает поднимание глотки и гортани, опускание мягкого неба и надгортанника при глотательном рефлексе. Чувствительное ядро одиночного пути получает вкусовую, тактильную, температурную, болевую и интероцептивную чувствительность от слизистой оболочки глотки, задней трети языка, барабанной полости и каротидного тельца, входит в состав соответствующих анализаторов, участвует в рефлексах жевания, глотания, в секреторных и моторных пищеварительных рефлексах, в сосудистых и сердечных рефлексах (из каротидного тельца). Нижнее слюноотделительное (парасимпатическое) ядро стимулирует секрецию околоушной слюнной железы.
пара X — блуждающий нерв; ядра блуждающего нерва расположены в продолговатом мозге. Двойное (двигательное) ядро, иннервируя мышцы неба, глотки, гортани, участвует в рефлекса глотания, рвоты, чиханья, кашля, в формировании голоса. Чувствительное ядро одиночного пути, иннервируя слизистую оболочку неба, корня языка, дыхательных путей, аортальное тельце, органы шеи, грудной, брюшной полостей, участвует в качестве афферентного звена в глотательном, жевательном дыхательных, висцеральных рефлексах. Оно входит в проводниковый отдел интероцептивного, вкусового, тактильного, температурного и болевого анализаторов. Заднее (парасимпатическое) ядро, иннервируя сердце, гладкие мышцы и железы органов шеи грудной и брюшной полостей, участвует сердечных, легочных, бронхиальных, пищи верительных рефлексах.
пара XI — добавочный нерв; двигательное ядро добавочного нерв расположено в продолговатом и спинном мозге. Иннервируя грудино-ключично-сосцевидную и трапециевидную мышцы, оно вызывает наклон головы набок с поворотом лица в противоположную сторону, поднимание плечевого пояса вверх приведение лопаток к позвоночнику.
пара XII — подъязычный нерв; двигательное ядро подъязычного нерва расположено в продолговатом мозге; иннервируя мышцы языка, вызывает его движение в рефлексах жевания, сосания, глотания, в осуществлении речи.
Таким образом, с участием ядер черепных нервов реализуется сенсорная и рефлекторная (соматическая и вегетативная) функции ствола мозга.
Сенсорные функции.Продолговатый мозг регулирует ряд сенсорных функций:
рецепцию кожной чувствительности лица — в сенсорном ядре тройничного нерва;
первичный анализ рецепции вкуса — в ядре языкоглоточного нерва;
рецепцию слуховых раздражений — в ядре улиткового нерва;
рецепцию вестибулярных раздражений — в верхнем вестибулярном ядре.
В задневерхних отделах продолговатого мозга проходят пути кожной, глубокой, висцеральной чувствительности, часть из которых переключается здесь на второй нейрон (тонкое и клиновидное ядра). На уровне продолговатого мозга перечисленные сенсорные функции реализуют первичный анализ силы и качества раздражения, далее обработанная информация передается в подкорковые структуры для определения биологической значимости данного раздражения.
Проводниковые функции. Через продолговатый мозг проходят все восходящие и нисходящие пути спинного мозга: спинно-таламический, кортикоспинальный, руброспинальный. В нем берут начало вестибулоспинальный, оливоспинальный и ретикулоспинальный тракты, обеспечивающие тонус и координацию мышечных реакций. В продолговатом мозге заканчиваются пути из коры большого мозга — корковоретикулярные пути. Здесь заканчиваются восходящие пути проприоцептивной чувствительности из спинного мозга:тонкого и клиновидного. Такие образования головного мозга, как мост, средний мозг, мозжечок, таламус, гипоталамус и кора большого мозга, имеют двусторонние связи с продолговатым мозгом.
Через восходящие пути и черепные нервы продолговатый мозг получает импульсы от рецепторов мышц лица, шеи, конечностей и туловища, от кожи лица, слизистых оболочек глаз, носовой и ротовой полости, от рецепторов слуха, вестибулярного аппарата, рецепторов гортани, трахеи, легких, интерорецепторов пищеварительного аппарата и сердечно-сосудистой системы.
Наличие этих связей свидетельствует об участии продолговатого мозга в регуляции тонуса скелетной мускулатуры, вегетативных и высших интегративных функций, анализе сенсорных раздражений.
Рефлекторные функции. В продолговатом мозге находятся центры как относительно простых, так и более сложных рефлексов. За счет продолговатого мозга осуществляются:
защитныерефлексы (мигание, слезоотделение, чиханье, кашлевой и рефлекс акта рвоты);
Эти рефлексы реализуются благодаря тому, что информация о раздражении рецепторов слизистой оболочки глаза, полости рта, гортани, носоглотки через чувствительные ветви тройничного и языкоглоточного нервов попадает в ядра продолговатого мозга. Отсюда идет команда к двигательным ядрам тройничного, блуждающего, лицевого, языкоглоточного, добавочного или подъязычного нервов, в результате реализуется тот или иной защитный рефлекс.
Точно так же за счет последовательного включения мышечных групп головы, шеи, грудной клетки и диафрагмы организуются рефлексы пищевого поведения: сосания, жевания, глотания.
установочныерефлексы, обеспечивающие тонус мускулатуры, необходимый для поддержания позы и осуществления рабочих актов;
Эти рефлексы формируются за счет афферентации от рецепторов преддверия улитки и полукружных каналов вверхнее вестибулярное ядро; отсюда переработанная информация оценки необходимости изменения позы посылается к латеральному и медиальному вестибулярным ядрам. Эти ядра участвуют в определении того, какие мышечные системы, сегменты спинного мозга должны принять участие в изменении позы, поэтому от нейроновмедиального и латерального ядра по вестибулоспинальному пути сигнал поступает к передним рогам соответствующих сегментов спинного мозга, иннервирующих мышцы, участие которых в изменении позы в данный момент необходимо.
лабиринтные рефлексы, способствующие правильному распределению мышечного тонуса между отдельными группами мышц при изменении позы тела;
Изменение позы осуществляется за счет статических и статокинетических рефлексов.Статические рефлексы регулируют тонус скелетных мышц с целью удержания определенного положения тела.Статокинетические рефлексы продолговатого мозга обеспечивают перераспределение тонуса мышц туловища для организации позы соответствующей моменту прямолинейного или вращательного движения..
рефлексы, связанные с функциями систем дыхания, кровообращения, пищеварения.
Большая часть автономных рефлексов продолговатого мозга реализуется через расположенные в нем ядра блуждающего нерва, которые получают информацию о состоянии деятельности сердца, сосудов, пищеварительного тракта, легких, пищеварительных желез и др. В ответ на эту информацию ядра организуют двигательную и секреторную реакции названных органов.
Возбуждение ядер блуждающего нерва вызывает усиление сокращения гладких мышц желудка, кишечника, желчного пузыря и одновременно расслабление сфинктеров этих органов. При этом замедляется и ослабляется работа сердца, сужается просвет бронхов.
Деятельность ядер блуждающего нерва проявляется также в усилении секреции бронхиальных, желудочных, кишечных желез, в возбуждении поджелудочной железы, секреторных клеток печени.
В продолговатом мозге локализуется центр слюноотделения, парасимпатическая часть которого обеспечивает усиление общей секреции, а симпатическая — белковой секреции слюнных желез.
Рефлекторные центры продолговатого мозга. В продолговатом мозге располагается ряд жизненно важных центров.В структуре ретикулярной формации продолговатого мозга расположены дыхательный и сосудодвигательный центры. Особенность этих центров в том, что их нейроны способны возбуждаться рефлекторно и под действием химических раздражителей.
Дыхательный центр локализуется в медиальной части ретикулярной формации каждой симметричной половины продолговатогомозга и разделен на две части, вдоха и выдоха.
Сосудодвигательный центр (регуляции сосудистого тонуса) функционирует совместно с вышележащими структурами мозга и прежде всего с гипоталамусом. Возбуждение сосудодвигательного центра всегда изменяет ритм дыхания, тонус бронхов, мышц кишечника, мочевого пузыря, цилиарной мышцы и др.
В средних отделах ретикулярной формации находятся нейроны, образующие ретикулоспинальный путь, оказывающий тормозное влияние на мотонейроны спинного мозга. На дне IV желудочка расположены нейроны «голубого пятна». Их медиатором является норадреналин. Эти нейроны вызывают активацию ретикулоспинального пути в фазу «быстрого» сна, что приводит к торможению спинальных рефлексов и снижению мышечного тонуса.
Средний мозг.
К образованиям среднего мозга относят ножки мозга, ядра III (глазодвигательный) и IV (блоковый) пар черепных нервов, четверохолмие, красные ядра и черное вещество (черную субстанцию). В ножках мозга проходят восходящие и нисходящие нервные пути.
В строении среднего мозга полностью утрачиваются сегментарные признаки. В среднем мозге клеточные элементы образуют сложные скопления в виде ядер. Ядерные образования относятся непосредственно к среднему мозгу, а также к входящей в его состав ретикулярной формации.
Передние бугры четверохолмияполучают импульсы от сетчатой оболочки глаз. В ответ на эти сигналы осуществляется регуляция просвета зрачка и аккомодация глаза.
Задние бугры четверохолмияполучают импульсы от ядер слуховых нервов, расположенных в продолговатом мозге. Благодаря этому происходит рефлекторная регуляция тонуса мышц среднего уха, а у животных - поворот ушной раковины к источнику звука. Таким образом, при участии передних и задних бугров четверохолмия осуществляютсяустановочные, ориентировочныерефлекторные реакции на световые и звуковые раздражения (движения глаз, поворот головы и даже туловища в сторону светового или звукового раздражителя). При разрушении ядер четверохолмия зрение и слух сохраняются, но отсутствуют ориентировочные реакции на свет и звук.
С деятельностью бугров четверохолмия тесно связана функция ядер IIIиIVпар черепных нервов, возбуждение которых определяет движение глаз вверх, вниз, в стороны, а также сведение (конвергенция) и разведение глазных осей при переносе взора с удаленных предметов на близкие и обратно.
Красные ядраучаствуют в регуляции мышечного тонуса и в проявлении установочных рефлексов, обеспечивающих сохранение правильного положения тела в пространстве. При отделении заднего мозга от среднего тонус мышц-разгибателей повышается, конечности животного напрягаются и вытягиваются, голова запрокидывается. Следовательно, у здорового животного и человека красные ядра несколько притормаживают тонус мышц-разгибателей.
Черное веществотакже регулирует мышечный тонус и поддержание позы, участвует в регуляции актов жевания, глотания, кровяного давления и дыхания, т. е. деятельность черного вещества, как и красных ядер, тесно связана с работой продолговатого мозга.
Таким образом, средний мозг регулирует тонус мышц, соответствующим образом его распределяет, что является необходимым условием координированных движений; Средний мозг регулирует ряд вегетативных функций организма (жевание, глотание, кровяное давление, дыхание). За счет среднего мозга расширяется, становятся многообразнее рефлекторная деятельность организма (ориентировочные рефлексы на звуковые и зрительные раздражения).
Тонические рефлексы ствола мозга
Образования ствола мозга обеспечивают правильное распределение тонуса между отдельными группами мышц. Рефлексы, обеспечивающие мышечный тонус, получили название тонических. В осуществлении этих рефлексов участвуют мотонейроны спинного мозга, вестибулярные ядра продолговатого мозга, мозжечок, образования среднего мозга (красные ядра). В целостном организме проявление тонических рефлексов контролируется клетками моторной зоны коры больших полушарий.
Тонические рефлексы возникают при изменении положения тела и головы в пространстве за счет возбуждения проприорецепторов и мышц, рецепторов вестибулярного аппарата внутреннего уха и тактильных рецепторов кожи.
Тонические рефлексы делят на две группы: статические и статокинетические. Статические рефлексы возникают при изменении положения тела, особенно головы, в пространстве. Статокинетические рефлексы проявляются при перемещении тела в пространстве, при изменении скорости движения (вращательного или прямолинейного).
Таким образом, тонические рефлексы предотвращают возможность нарушения равновесия, потерю активной позы и способствуют восстановлению нарушенной позы.
Промежуточный мозг.
Промежуточный мозг - часть переднего отдела ствола мозга. Основными образованиями промежуточного мозга являются зрительные бугры (таламус) и подбугорная область (гипоталамус).
Зрительные бугры-массивное парное образование, они занимают основную массу промежуточного мозга. Наибольших размеров и наивысшей сложности строения зрительные бугры достигают у человека.
Зрительные бугры являются центром всех афферентных импульсов. Через зрительные бугры к коре головного мозга поступает информация от всех рецепторов нашего организма, за исключением обонятельных. Кроме того, от зрительных бугров нервные импульсы передаются к различным образованиям ствола мозга. В зрительных буграх обнаружено большое количество ядерных образований. Функционально их можно разделить на две группы: специфические и неспецифические ядра.
Специфические ядраполучают информацию от рецепторов, перерабатывают ее и передают в определенные области коры головного мозга, где возникают соответствующие ощущения (зрительные, слуховые и т. д.).Неспецифические ядране имеют прямой связи с рецепторами организма. Они получают импульсы от рецепторов через большое количество переключений (синапсов). Импульсы от этих образований через подкорковые ядра поступают к множеству нейронов, расположенных в различных областях коры головного мозга, вызывая повышение их возбудимости.
При повреждении зрительных бугров у человека наблюдается полная потеря чувствительности или ее снижение на противоположной стороне, выпадает сокращение мимической мускулатуры, которое сопровождает эмоции, также могут возникать расстройства сна, понижение слуха, зрения и т. д.
Гипоталамическая (подбугорная) областьучаствует в регуляции различных видов обмена веществ (белков, жиров, углеводов, солей, воды), регулирует теплообразование и теплоотдачу, состояние сна и бодрствования; В ядрах гипоталамуса происходит образование ряда гормонов, которые затем депонируются в задней доле гипофиза.. Передние отделы гипоталамуса являются высшими центрами парасимпатической нервной системы, задние - симпатической нервной системы. Гипоталамус участвует в регуляции многих вегетативных функций организма.
Базальные ядра.
К подкорковым, или базальным, ядрам относятся три парных образования: хвостатое ядро, скорлупа и бледный шар. Базальные ядра расположены внутри больших полушарий, в нижней их части, между лобными долями и промежуточным мозгом. Развитие и клеточное строение у хвостатого ядра и скорлупы одинаковы, поэтому их рассматривают как единое образование -полосатое тело.
Полосатое теловедает сложными двигательными функциями, участвует в осуществлении безусловнорефлекторных реакций цепного характера - бег, плавание, прыжки. Эту функцию полосатое тело осуществляет через бледный шар, притормаживая его деятельность. Кроме того, полосатое тело через гипоталамус регулирует вегетативные функции организма, а также вместе с ядрами промежуточного мозга обеспечивает осуществление сложных безусловных рефлексов цепного характера -инстинктов.
Бледный шарявляется центром сложных двигательных рефлекторных реакций (ходьба, бег), формирует сложные мимические реакции, участвует в обеспечении правильного. распределения мышечного тонуса. Свои функции бледный шар осуществляет опосредованно через образования среднего мозга (красные ядра и черное вещество). При раздражении бледного шара наблюдается общее сокращение скелетных мышц противоположной стороны тела. При поражении бледного шара движения теряют свою плавность, становятся неуклюжими, скованными.
Следовательно, деятельность подкорковых ядер не ограничивается их участием в формировании сложных двигательных актов. Они благодаря связям с гипоталамусом участвуют в регуляции обмена веществ и функций внутренних органов.
Таким образом, базальные ядра являются высшими подкорковыми центрами объединения (интеграции) функций организма. У человека и высших позвоночных животных деятельность подкорковых ядер контролируется корой головного мозга.
Ретикулярная формация ствола мозга
Ретикулярная формация представляет собой скопление особых нейронов, которые своими волокнами образуют своеобразную сеть. Нейроны ретикулярной формации в области ствола мозга были описаны в прошлом веке немецким ученым Дейтерсом. В. М. Бехтерев подобные же структуры обнаружил в области спинного мозга. Нейроны ретикулярной формации образуют скопления, или ядра. Дендриты этих клеток относительно длинные, мало ветвистые, аксоны, напротив, короткие, имеют много ответвлений (коллатералей). Эта особенность обусловливает многочисленные синаптические контакты нейронов ретикулярной формации.
Ретикулярная формация ствола мозга занимает центральное положение в продолговатом мозге, варолиевом мосту, среднем и промежуточном мозге.
Нейроны ретикулярной формации не имеют непосредственных контактов с рецепторами организма. Нервные импульсы при возбуждении рецепторов поступают к ретикулярной формации по коллатералям волокон вегетативной и соматической нервной системы.
Физиологическая роль. Ретикулярная формация ствола мозга оказывает восходящее влияние на клетки коры головного мозга и нисходящее на мотонейроны спинного мозга. Оба эти влияния ретикулярной формации могут быть активирующими или тормозными.
Афферентная импульсация к коре головного мозга поступает по двум путям: специфическому и неспецифическому. Специфический нервный путьобязательно проходит через зрительные бугры и несет нервные импульсы к определенным зонам коры головного мозга, в результате осуществляется какая-либо специфическая деятельность. Например, при раздражении фоторецепторов глаз импульсы через зрительные бугры поступают в затылочную область коры головного мозга и у человека возникают зрительные ощущения.
Неспецифический нервный путьобязательно проходит через нейроны ретикулярной формации ствола мозга. Импульсы к ретикулярной формации поступают по коллатералям специфического нервного пути. Благодаря многочисленным синапсам на одном и том же нейроне ретикулярной формации могут сходиться (конвергировать) импульсы различных значений (световые, звуковые и т. д.), при этом они теряют свою специфичность. От нейронов ретикулярной формации эти импульсы поступают не в какую-то определенную область коры головного мозга, а веерообразно распространяются по ее клеткам, повышая их возбудимость и облегчая тем самым выполнение специфической функции.
В опытах на кошках с вживленными в область ретикулярной формации ствола мозга электродами было показано, что раздражение ее нейронов вызывает пробуждение спящего животного. При разрушении ретикулярной формации животное впадает в длительное сонное состояние. Эти данные свидетельствуют о важной роли ретикулярной формации в регуляции состояния сна и бодрствования. Ретикулярная формация не только оказывает влияние на кору головного мозга, но также посылает в спинной мозг к его двигательным нейронам тормозящие и возбуждающие импульсы. Благодаря этому она участвует в регуляции тонуса скелетной мускулатуры.
В спинном мозге, как уже указывалось, также имеются нейроны ретикулярной формации. Полагают, что они поддерживают на высоком уровне активность нейронов спинного мозга. Функциональное состояние самой ретикулярной формации регулируется корой головного мозга.
Мозжечок
Особенности строения мозжечка.Связи мозжечка с другими отделами центральной нервной системы. Мозжечок - это непарное образование; он располагается позади продолговатого мозга и варолиева моста, граничит с четверохолмиями, сверху прикрыт затылочными долями больших полушарий, В мозжечке различают среднюю часть -червьи расположенные по бокам от него дваполушария. Поверхность мозжечка состоит изсерого вещества,называемого корой, которая включает тела нервных клеток. Внутри мозжечка располагаетсябелое вещество, представляющее собой отростки этих нейронов.
Мозжечок имеет обширные связи с различными отделами центральной нервной системы за счет трех пар ножек. Нижние ножкисоединяют мозжечок со спинным и продолговатым мозгом,средние- с варолиевым мостом и через него с двигательной областью коры головного мозга,верхние-со средним мозгом и гипоталамусом.
Функции мозжечка были изучены на животных, у которых мозжечок удаляли частично или полностью, а также путем регистрации его биоэлектрической активности в покое и при раздражении.
При удалении половины мозжечка отмечается повышение тонуса мышц-разгибателей, поэтому конечности животного вытягиваются, наблюдаются изгиб туловища и отклонение головы в оперированную сторону, иногда качательные движения головой. Часто движения совершаются по кругу в оперированную сторону («манежные движения»). Постепенно отмеченные нарушения сглаживаются, однако сохраняется некоторая неловкость движений.
При удалении всего мозжечка наступают более выраженные двигательные расстройства. В первые дни после операции животное лежит неподвижно с запрокинутой головой и вытянутыми конечностями. Постепенно тонус мышц-разгибателей ослабевает, появляется дрожание мышц, особенно шейных. В дальнейшем двигательные функции частично восстанавливаются. Однако до конца жизни животное остается двигательным инвалидом: при ходьбе такие животные широко расставляют конечности, высоко поднимают лапы, т. е. у них нарушена координация движений.
Двигательные расстройства при удалении мозжечка были описаны известным итальянским физиологом Лючиани. Основными из них являются: а т о н и я - исчезновение или ослабление мышечного тонуса; а с т е н и я -снижение силы мышечных сокращений. Для такого животного характерно быстро наступающее мышечное утомление; а с т а з и я - потеря способности к слитным тетаническим сокращениям, У животных наблюдаются дрожательные движения конечностей и головы. Собака после удаления мозжечка не может сразу поднять лапы, животное делает ряд колебательных движений лапой, перед тем как ее поднять. Если поставить такую собаку, то тело ее и голова все время качаются из стороны в сторону.
В результате атонии, астении и астазии у животного нарушается координация движений: отмечаются шаткая походка, размашистые, неловкие, неточные движения. Весь комплекс двигательных расстройств при поражении мозжечка получил название мозжечковой атаксии.
Подобные нарушения наблюдаются и у человека при поражении мозжечка.
Через некоторое время после удаления мозжечка, как уже указывалось, все двигательные расстройства постепенно сглаживаются. Если у таких животных удалить моторную область коры головного мозга, то двигательные нарушения вновь усиливаются. Следовательно, компенсация (восстановление) двигательных расстройств при поражении мозжечка осуществляется при участии коры головного мозга, ее моторной области.
Исследованиями Л. А. Орбели было показано, что при удалении мозжечка наблюдается не только падение мышечного тонуса (атония), но и неправильное его распределение (дистония). Л. Л. Орбели установил, что мозжечок влияет и на состояние рецепторного аппарата, а также на вегетативные процессы. Мозжечок оказывает адаптационно-трофическое влияние на все отделы мозга через симпатическую нервную систему, он регулирует обмен веществ в головном мозге и тем самым способствует приспособлению нервной системы к изменяющимся условиям существования.
Таким образом, основными функциями мозжечка являются координация движений, нормальное распределение мышечного тонуса и регуляция вегетативных функций. Свое влияние мозжечок реализует через ядерные образования среднего и продолговатого мозга, через двигательные нейроны спинного мозга. Большая роль в этом влиянии принадлежит двусторонней связи мозжечка с моторной зоной коры головного мозга и ретикулярной формацией ствола мозга.
Особенности строения коры больших полушарий головного мозга.
Кора больших полушарий головного мозга в филогенетическом отношении является высшим и наиболее молодым отделом центральной нервной системы.
Кора мозга состоит из нервных клеток, их отростков и нейроглии. У взрослого человека толщина коры в большинстве областей составляет около 3 мм. Площадь корыбольших полушарий благодаря многочисленным складкам и бороздам составляет 2500 см2. Для большинства участков коры головного мозга характерно шестислойное расположение нейронов. Кора больших полушарий состоит из 14—17 млрд. клеток. Клеточные структуры коры головного мозга представлены пирамидными, веретенообразными и звездчатыми нейронами.
Звездчатые клеткивыполняют главным образом афферентную функцию. Пирамидные и веретенообразные клетки — это преимущественно эфферентные нейроны.
В коре больших полушарий имеются высокоспециализированные нервные клетки, воспринимающие афферентные импульсы от определенных рецепторов (например, от зрительных, слуховых, тактильных и т. д.). Имеютсятакже нейроны, которые возбуждаются нервными импульсами, идущими от разных рецепторов организма. Это так называемые полисенсорные нейроны.
Отростки нервных клеток коры головного мозга связывают ее различные отделы между собой или устанавливают контакты коры больших полушарий с нижележащими отделами центральной нервной системы. Отростки нервных клеток, соединяющие между собой различные участки одного и того же полушария называются ассоциативными, связывающие чаще всего одинаковые участки двух полушарий — комиссуральными и обеспечивающие контакты коры головного мозга с другими отделами центральной нервной системы и через них со всеми органами и тканями тела — проводящими (центробежными). Схема этих путей приведена на рисунке.
Схема хода нервных волокон в больших полушариях головного мозга.
1 — короткие ассоциативные волокна; 2 — длинные ассоциативные волокна; 3 - комиссуральные волокна; 4 — центробежные волокна.
Клетки нейроглии выполняют ряд важных функций: они являются опорной тканью, участвуют в обмене веществ головного мозга, регулируют кровоток внутри мозга, выделяют нейросекрет, который регулирует возбудимость нейронов коры головного мозга.
Функции коры головного мозга.
1) Кора головного мозга осуществляет взаимодействие организма с окружающей средой за счет безусловных и условных рефлексов;
2) она является основой высшей нервной деятельности (поведения) организма;
3) за счет деятельности коры головного мозга осуществляются высшие психические функции: мышление и сознание;
4) кора головного мозга регулирует и объединяет работу всех внутренних органов и регулирует такие интимные процессы, как обмен веществ.
Таким образом, с появлением коры головного мозга она начинает контролировать все процессы, протекающие в организме, а также всю деятельность человека,т. е. происходит кортиколизация функций. И. П. Павлов, характеризуя значение коры головного мозга, указывал, что она является распорядителем и распределителем всей деятельности животного и человеческого организма.
Функциональное значение различных областей коры головного мозга. Локализация функций в коре головного мозга. Роль отдельных областей коры головного мозга впервые была изучена в 1870 г. немецкими исследователями Фричем и Гитцигом. Они показали, что раздражение различных участков передней центральной извилины и собственно лобных долей вызывает сокращение определенных групп мышц на противоположной раздражению стороне. В дальнейшем была выявлена функциональная неоднозначность различных областей коры. Было обнаружено, что височные доли коры головного мозга связаны со слуховыми функциями, затылочные — со зрительными функциями и т.д. Эти исследования позволили сделать вывод, что разные участки коры больших полушарий ведают определенными функциями. Было создано учение о локализации функций в коре головного мозга.
По современным представлениям, различают три типа зон коры головного мозга: первичные проекционные зоны, вторичные и третичные (ассоциативные).
Первичные проекционные зоны — это центральные отделы ядер анализаторов. В них расположены высокодифференцированные и специализированные нервные клетки, к которым поступают импульсы от определенных рецепторов (зрительных, слуховых, обонятельных и др.). В этих зонах происходит тонкий анализ афферентных импульсов различного значения. Поражение указанных зон ведет к расстройствам чувствительных или двигательных функций.
Вторичные зоны — периферические отделы ядер анализаторов. Здесь происходит дальнейшая обработка информации, устанавливаются связи между различными похарактеру раздражителями. При поражении вторичных зон возникают сложные расстройства восприятий.
Третичные зоны (ассоциативные). Нейроны этих зон могут возбуждаться под влиянием импульсов, идущих от рецепторов различного значения (от рецепторов слуха, фоторецепторов, рецепторов кожи и т. д.). Это так называемые полисенсорные нейроны, за счет которых устанавливаются связи между различными анализаторами. Ассоциативные зоны получают переработанную информацию от первичных и вторичных зон коры больших полушарий. Третичные зоны играют большую роль в формировании условных рефлексов, они обеспечивают сложные формы познания окружающей действительности.
Значение различных областей коры головного мозга. В коре большого мозга выделяют сенсорные, моторные области
Сенсорные области коры. (проекционная кора, корковые отделы анализаторов). Это зоны, в которые проецируются сенсорные раздражители. Они расположены преимущественно в теменной, височной и затылочной долях. Афферентные пути в сенсорную кору поступают преимущественно от релейных сенсорных ядер таламуса – вентральных задних, латерального и медиального. Сенсорные области коры образованы проекционными и ассоциативными зонами основных анализаторов.
Область кожной рецепции (мозговой конец кожного анализатора) представлена в основном задней центральной извилиной. Клетки этой области воспринимают импульсы от тактильных, болевых и температурных рецепторов кожи. Проекция кожной чувствительности в пределах задней центральной извилины аналогична таковой для двигательной зоны. Верхние участки задней центральной извилины связаны с рецепторами кожи нижних конечностей, средние — с рецепторами туловища и рук, нижние — с рецепторами кожи головы и лица. Раздражение этой .области у человека во время нейрохирургических операций вызывает ощущения прикосновения, покалывания, онемения, при этом никогда не наблюдается выраженных болевых ощущений.
Область зрительной рецепции (мозговой конец зрительного анализатора) расположена в.затылочных долях коры головного мозга обоих полушарий. Эту область следует рассматривать как проекцию сетчатой оболочки глаза.
Область слуховой рецепции (мозговой конец слухового анализатора) локализуется в височных долях коры головного мозга. Сюда поступают нервные импульсы от рецепторов улитки внутреннего уха. При повреждении этой зоны может возникнуть музыкальная и словесная глухота, когда человек слышит, но не понимает значения слов; Двустороннее поражение слуховой области приводит к полной глухоте.
Область вкусовой рецепции (мозговой конец вкусового анализатора) расположена в нижних долях центральной извилины. Эта область получает нервные импульсы от вкусовых рецепторов слизистой оболочки полости рта.
Область обонятельной рецепции (мозговой конец обонятельного анализатора) располагается в передней части грушевидной доли коры головного мозга. Сюда поступают нервные импульсы от обонятельных рецепторов слизистой оболочки носа.
В коре больших полушарий обнаружено несколько зон, ведающих функцией речи (мозговой конец речедвигательного анализатора). В лобной области левого полушария (у праворуких) располагается моторный центр речи (центр Брока). При его поражении речь затруднена или даже невозможна. В височной области находится сенсорный центр речи (центр Вернике). Повреждение этой области приводит к расстройствам восприятия речи: больной не понимает значение слов, хотя способность произносить слова сохранена. В затылочной доле коры головного Мозга имеются зоны, обеспечивающие восприятие письменной (зрительной) речи. При поражении этих областей больной не понимает написанного.
В теменной области коры больших полушарий не обнаружены мозговые концы анализаторов, ее относят к ассоциативным зонам. Среди нервных клеток теменной области найдено большое количество полисенсорных нейронов, которые способствуют установлению связей между различными анализаторами и играют большую роль в формировании рефлекторных дуг условных рефлексов
Моторные области корыПредставление о роли двигательной коры большого мозга двояко. С одной стороны, было показано, что электрическое раздражение некоторых корковых зон у животных вызывает движение конечностей противоположной стороны тела, что говорило о том, что кора непосредственно участвует в реализации двигательных функций. В то же время признано, что двигательная область является анализаторной, т.е. представляет собой корковый отдел двигательного анализатора.
Мозговой отдел двигательного анализатора представлен передней центральной извилиной и расположенными вблизи нее участками лобной области. При ее раздражении возникают разнообразные сокращения скелетной мускулатуры на противоположной стороне. Установлено соответствие между определенными зонами передней центральной извилины и скелетной мускулатурой. В верхних участках этой зоны проецируется мускулатура ног, в средних — туловища, в нижних — головы.
Особый интерес представляет собственно лобная область, которая достигает у человека наибольшего развития. При поражении лобных областей у человека нарушаются сложные двигательные функции, обеспечивающие трудовую деятельность и речь, а также приспособительные, поведенческие реакции организма.
Любая функциональная зона коры головного мозга находится и в анатомическом, и в функциональном контакте с другими зонами коры больших полушарий, с подкорковыми ядрами, с образованиями промежуточного мозга и ретикулярной формации, что обеспечивает совершенство выполняемых ими функций.
Профильный материал для студентов педиатрического факультета
1. Структурно-функциональные особенности цнс в антенатальном периоде.
- У плода количество нейронов ДНС достигает максимума к 20—24-й неделе и остается в постнатальном периоде без резкого снижения до пожилого возраста. Нейроны имеют малые размеры и суммарную площадь синаптической мембраны.
- Аксоны развиваются раньше дендритов, отростки нейронов интенсивно растут и ветвятся. Наблюдается увеличение длины, диаметра и миелинизации аксонов к концу антенатального периода.
- Филогенетически старые пути миелинизируются раньше, чем филогенетически новые; например, вестибулоспинал ьные пути с 4-го месяца внугриугробного развития, руброспинальные пути с 5—8-го месяца, пирамидные пути после рождения.
- Nа- и К-каналы равномерно распределены в мембране миелиновых и немиелиновых волокон.
- Возбудимость, проводимость, лабильность нервных волокон значительно ниже, чем у взрослых людей.
- Синтез большинства медиаторов начинается в период внутриутробного развития. Гамма-аминомасляная кислота в антенатальном периоде является возбуждающим медиатором и через Са2-механизм оказывает морфогенные эффекты — ускоряет рост аксонов и дендрвтов, синаптогенез, экспрессию питорецепторов.
- К моменту рождения заканчивается процесс дифференциации нейронов ядер продолговатого и среднего мозга, моста.
- Имеется структурно-функциональная незрелость глиальных клеток.
2. Особенности цнс в периоде новорожденности.
> Возрастает степень миелинизации нервных волокон, их количество составляет 1/з уровня взрослого организма (например, полностью миелинизирован руброспинальный путь).
> Уменьшается проницаемость клеточных мембран для ионов. Нейроны имеют более низкую амплитуду МП — около 50 мВ (у взрослых примерно 70 мВ).
> На нейронах синапсов меньше, чем у взрослых, мембрана нейрона имеет рецепторы к синтезируемым медиаторам (ацетилхолину, ГАМ К, серотонину, норадреналину в дофамину). Содержание медиаторов в нейронах мозга новорожденных низкое в составляет 10—50% медиаторов у взрослых.
> Отмечается развитие шипикового аппарата нейронов и аксошипиковых синапсов; ВПСП и ТПСП имеют большую длительность и меньшую амплитуду, чем у взрослых. Количество тормозных синапсов на нейронах меньше, чем у взрослых.
> Повышается возбудимость корковых нейронов.
> Исчезает (точнее, резко уменьшается) митотическая активность и возможность регенерации нейронов. Продолжается пролиферация и функциональное созревание глиоцитов.
З. Особенности ЦНС в грудном возрасте.
- Созревание ЦНС быстро прогрессирует. Наиболее интенсивная миелинизация нейронов ЦНС происходит в конце первого года после рождения (например, к 6 мес завершается миелинизация нервных волокон полушарий мозжечка).
- Возрастает скорость проведения возбуждения по аксонам.
- Наблюдается уменьшение продолжительности ПД нейронов, укорачиваются абсолютная и относительная рефрактерные фазы (длительность абсолютной рефрактерности 5—8 мс, относительной 4О-бО мс в раннем постнатальном онтогенезе, у взрослых соответственно 0,5—2,О и 2—10 мс).
- Кровоснабжение мозга у детей относительно больше, чем у взрослых.