

Электропорация.
Электропорация была проведена, например, для эмбрионов креветки и моллюсков. Недостатком использования электропорации является постепенная деградация и уменьшение экспрессии трансгена со временем.
Биобаллистическая трансформация
Суть метода биобаллистической трансформации заключается в том, что на мельчайшие частички вольфрама, платины или золота, диаметром от 0,1 до 3,5 мкм, напыляется векторная ДНК, содержащая необходимую для трансформации генную конструкцию. Вольфрамовые, платиновые или золотые частички, несущие ДНК, на целлофановой подложке помещаются внутрь биобаллистической пушки. Суспензия животных клеток или эмбрионов, на ранней стадии развития, помещается под биобаллистическую пушку на расстоянии 10-25 см. В пушке вакуумным насосом уменьшается давление до 0,1 атм. В момент сбрасывания давления частички металла с огромной скоростью выбрасываются из пушки и, пробивая мембраны, входят в цитоплазму и ядра клеток. Обычно клетки, располагающиеся непосредственно по центру, погибают из-за огромного количества и давления частичек металла, в то время как в зоне 0,6-1 см от центра будут находиться трансформированные клетки (Рис.3). Далее клетки или эмбрионы животных переносят на среду для дальнейшего культивирования и регенерации. Бомбардировка микрочастицами была использована, например, для трансформации оплодотворенных яйцеклеток креветки, морского ежа. Процент выживших клеток после бомбардировки составляет приблизительно 70%, а также была доказана экспрессия трансгена у некоторых разновидностей рыб. Главным преимуществом данного метода является высокая эффективность встройки векторной ДНК, а также то, что можно получить трансгенные клетки в самые кратчайшие сроки.
Биобаллистическая трансформация.
Существенным недостатком этого метода является то, что эмбрионы, подвергшиеся бомбардировке, довольно редко развиваются в полноценных взрослых особей, большая часть из них рано или поздно гибнет.
Липофекция Липосомы - это сферические образования, оболочки которых состоят из фосфолипидов. Их можно получить в результате резкого встряхивания или обработки ультразвуком водных эмульсий фосфолипидов. Метод липофекции основан на взаимодействии между положительно заряженными молекулами фосфолипидов, из которых состоят липосомы, и отрицательно заряженными молекулами ДНК. В настоящее время предложены три модели ассоциаций между ДНК и фосфолипидами . Суть первой модели состоит в том, что положительно заряженные липосомы присоединяются к отрицательно заряженным молекулам ДНК. Число липосом, присоединившихся к ДНК, зависит от размеров молекулы нуклеиновой кислоты. Во втором случае молекулы ДНК "проглатываются" липосомами, то есть ДНК электростатически взаимодействует с внутренней поверхностью липосомы. В третьем случае молекула ДНК окружена несколькими молекулами фосфолипидов, которые формируют своеобразную ленту с нуклеиновой кислотой.
Липофекция (2-я модель).
Липосомы, несущие положительный заряд, легко присоединяются к несущей отрицательный заряд плазматической мембране животных клеток, после чего путем эндоцитоза проникают в цитоплазму клеток. Механизмы же, обеспечивающие встраивание в геном и экспрессию ДНК, не достаточно изучены. К преимуществам данного метода можно отнести низкую токсичность липосом по отношению к клеткам, а также то, что экзогенный генетический материал защищен от действия нуклеаз посредством транспортировки в липосомах. Недостатком данного метода является то, что количество личинок, экспрессирующих трансген сокращается с возрастом.
Более простой способ идентификации ЕS-клеток, несущих трансген в нужном сайте, основан на использовании ПЦР. В этом случае ДНК-вектор содержит два участка, гомологичных сайту-мишени, по одному со стороны трансгена и со стороны клонированной бактериальной или синтетической (уникальной) последовательности, отсутствующей в геноме мыши (рис. 6). После трансфекции ЕS-клеток этим вектором проводят скрининг трансфицированных клеток методом ПЦР. Один из ПЦР-праймеров (Р1) комплементарен участку клонированной бактериальной или синтетической (уникальной) нуклеотидной последовательности интегрировавшего вектора, а второй (Р2) — участку хромосомной ДНК, прилегающему к одному из гомологичных участков ДНК. При встраивании последовательности-мишени в случайный сайт ожидаемый продукт амплификации образовываться не будет (рис. 6, А), а при сайт-специфической интеграции в результате ПЦР-амплификации образуется фрагмент ДНК известного размера (рис. 6, Б), Таким образом можно идентифицировать пулы ЕS-клеток, содержащих трансген в нужном сайте, а пересевая клетки из этих пулов — получить клеточные линии с сайт-специфической вставкой.
ЕS-клетки, в геном которых в нужном сайте встроен трансген, можно культивировать и ввести в эмбрион на стадии бластоцисты, а затем имплантировать такие эмбрионы в матку псевдобеременных «суррогатных» матерей. Мышата, у которых генетически модифицированные ЕS-клетки участвовали в образовании клеток зародышевой линии, могут дать начало трансгенным линиям. Для этого их нужно скрестить с особями той же линии, а затем скрестить их трансгенных потомков. В результате будут получены трансгенные мыши, гомозиготные по трансгену.
В специфический хромосомный сайт ЕS-клеток можно не только встроить трансген, кодирующий какую-то новую функцию, но и направленно разрушить этот сайт интеграцией с его кодирующей областью специфической последовательности (обычно селективного маркерного гена) (рис. 7). Одна из задач направленного нарушения («нокаута») гена состоит в исследовании влияния этого процесса на развитие организма и протекающие в нем физиологические процессы. Кроме того, есть надежда, что трансгенных животных с нарушением в определенном гене можно использовать как модель для изучения болезней человека на молекулярном уровне.
Например, направленный «нокаут» гена родопсина мыши приводит к инактивации палочек сетчатки, что имитирует такую болезнь человека, как пигментный ретинит. На мышах с «нокаутированным» геном родопсина можно изучать процесс дегенерации сетчатки, а также терапевтический эффект лекарственных средств, замедляющих или вообще останавливающих генетически обусловленный патологический процесс. Уже создано более 250 линий мышей с «нокаутированными» генами, используюшихся в качестве моделей для изучения различных заболеваний человека.
В принципе подходы к созданию трансгенных животных с «улучшенными функциями» и с «потерей функций» сходны. К сожалению, плюрипотентные ЕS-клетки, аналогичные таковым у мышей, пока не обнаружены у крупного рогатого скота, овей, свиней и цыплят, но их поиск продолжается.
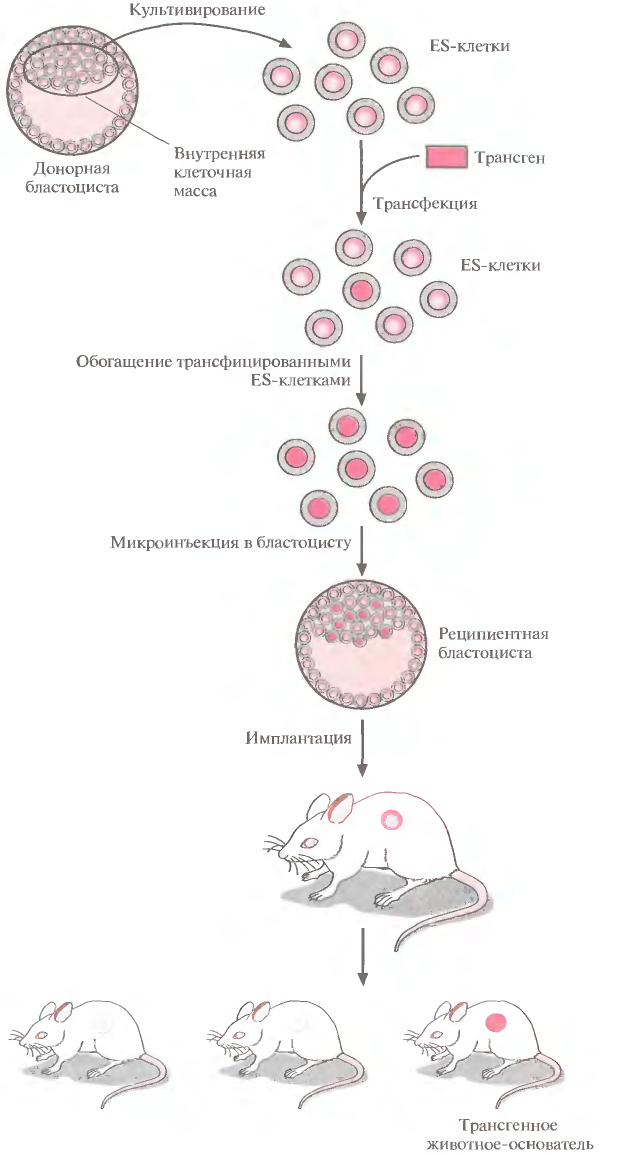
Рис. 4. Получение трансгенных мышей с помощью генетической модификации эмбриональных стволовых клеток.
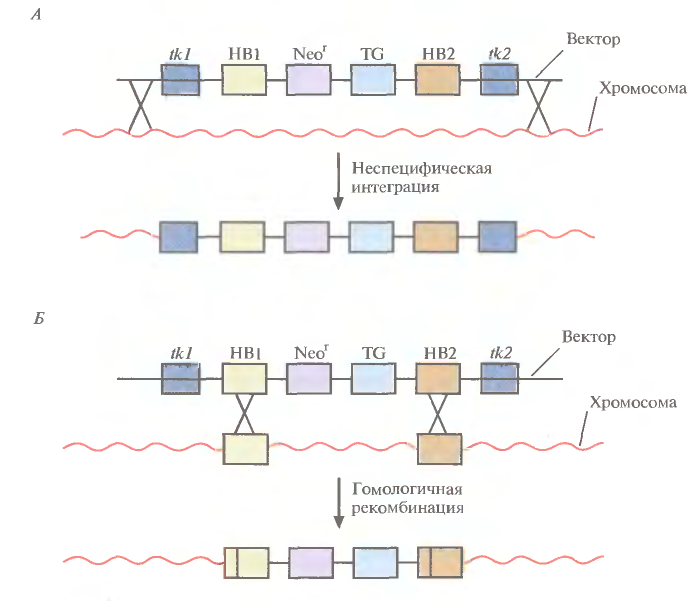
Рис. 5. Позитивно-негативная селекция. А. Неспецифическая интеграция. В хромосому встроились оба гена тимидинкиназы (tk1 и tk2), два участка ДНК, гомологичные специфичным последовательностям хромосомной ДНК реципиентных клеток (НВ1 и НВ2), ген (Neor), обеспечивающий устойчивость к цитотоксическому соединению G-418, и трансген (ТG). После трансфекции проводят тестирование клеток на устойчивость к G-418 и ганцикловиру, который становится цититоксичным для клеток, синтезирующих тимидинкиназу. Интеграция может произойти и по-другому, со встраиванием в хромосому только гена тимидинкиназы. В присутствии G-418 и ганцикловира все такие клетки тоже погибают. Б. Специфическая интеграция с помощью гомологичной рекомбинации. В результате двойного кроссинговера между гомологичными участками (НВ1 и HB2) векторной и хромосомной ДНК в последнюю встраивается фрагмент, не содержащий генов тимидинкиназы (tk1 и tk2). В присутствии G-418 и ганцикловира выживают только клетки, в которых прошла гомологичная рекомбинация.

Рис. 6. Идентификация клеток, несущих трансген в специфическом сайте, при помощи ПЦР. А. В результате неспецифического встраивания векторной ДНК один из праймеров (Р2) не сможет гибридизоваться с участком хромосомы, находящимся на определенном расстоянии от места отжига праймера Р1, и фрагмента нужного размера при амплификации не образуется. Р1 гибридизуется с уникальным участком (US) встроенной ДНК, отсутствующим в хромосомной ДНК клетки-реципнента. Б. В результате гомологичной рекомбинации между участками НВ1 и НВ2 встраиваемой ДНК, с одной стороны, и комплементарными участками хромосомы СS1 и СS2, с другой, образуются участки, с которыми могут гибридизоваться оба праймера, Р1 и Р2, и которые находятся на определенном расстоянии друг от друга. В ходе ПЦР-амплификации синтезируются фрагменты одного размера, которые можно идентифицировать при помощи гель-электрофореза. Если ПЦР-продукт нужной длины образовался, значит трансген (ТG), находящийся между гомологичными участками (НВ1 и НВ2), встроился в определенный сайт хромосомы.
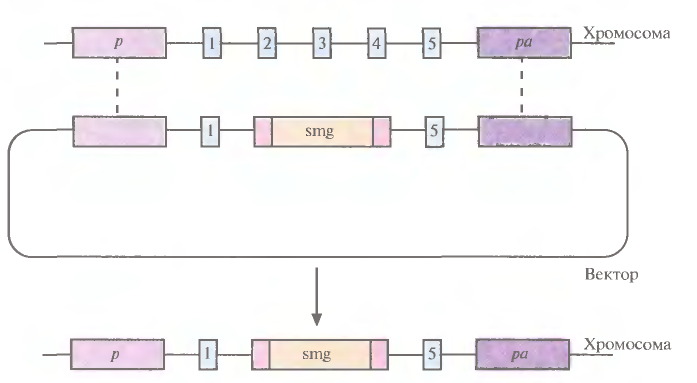
Рис. 7. «Нокаут» гена с помощью направленной гомологичной рекомбинации. Вектор несет селективный маркерный ген (smg) и фланкирующие его последовательности, гомологичные соответствующим участкам гена-мишени. Последний содержит пять экзонов (1 – 5). В результате гомологичной рекомбинации (штриховые линии) ген-мишень прерывается («нокаутируется»).