

Питання для самоконтролю
Як визначають оптимальний час для осіменіння корів та телиць?
Які методи застосовують для штучного осіменіння корів та телиць?
В чому суть цервікального осіменіння корів з ректальною фіксацією шийки матки?
Які особливості мано-цервікального осіменіння корів?
Як проводять візо-цервікальне осіменіння корів та телиць?
Як виявляють овець у стані охоти?
". Якими методами користуються при штучному осіменінні овець?
І. Як визначають оптимальний час для осіменіння свиноматок?
9. Якими методами штучного осіменіння користуються у свинарстві?
. Як визначають оптимальний час для осіменіння кобил?
187
6
ЗАПЛІДНЕННЯ
zygotos
Запліднення - складний біологічний процес, що полягає у проникненні спермія у цитоплазму яйцеклітини, об'єднанні їх ядер з утворенням нового типу клітини -зиготи (гр. гу§оіо8 - з'єднані разом), з якої шляхом відповідної трансформації в геніталіях самки розвивається плід з властивими йому індивідуальними особливостями росту, розвитку та продуктивності.
Пересування сперміїв у статевих органах самки. Запліднення відбувається у верхній третині яйцепроводу, у якому виділяють лійку, ампулу, перешийок (істмус) та інтрамуральний відділ. Перешийок є місцем природного резервування сперміїв, а ампула - місцем запліднення. Введені у статеві шляхи самки спермії для участі у заплідненні повинні пройти у ампулоподібне розширення яйцепроводів. Яким чином вони долають цю відстань?
Ще з часів А. Левенгука, коли йому вперше вдалося побачити під мікроскопом рухливих сперміїв, у науці тривалий час існувала думка, що назустріч яйцеклітині спермії просуваються виключно за рахунок власного руху. Проте з часом ця думка змінилася.
У процесі еволюції у ссавців виробилися пристосування, що сприяють зустрічі чоловічих та жіночих статевих клітин. Це, перш за все, відповідна морфологія та фізіологія статевої системи, морфологія та фізіологія гамет. Якщо, наприклад, співставити рух сперміїв тварин із зовнішнім і внутрішнім осіменінням, то у перших нормальним є коловий рух, оскільки при прямолінійному русі вони будуть розходитися у різні боки від місця знаходження яйцеклітини, тоді як у других саме прямолінійний поступальний рух буде сприяти переміщенню сперміїв від місця введення до місця запліднення.
Проходженню сперміїв уздовж статевих шляхів самки сприяє їх здатність до ре-отаксису - руху проти течії. Це було доведено експериментально: нанесені на розтягнену поверхню розрізаного яйцепроводу спермії рухалися по слизовій оболонці, долаючи успішно опір війок та течії рідини.
Осіменіння свійських тварин проводиться під час тічки, виділювані при цьому секрети статевих органів самки рухаються у протилежному напрямку і сприяють ре-отаксису сперміїв, як у піхві, так і у яйцепроводах.
Проте одним реотаксисом не можна пояснити швидкого проникнення сперміїв у яйцепроводи. У тварин з матковим типом осіменіння цьому сприяють особливос-
188
Запліднення
ті статевого акту. Великий об'єм еякуляту, заповнюючи просвіт рогів матки, сприяє швидкому наближенню сперміїв до яйцепроводу. У тварин з піхвовим типом осіменіння з великої кількості введених сперміїв (4-5 млрд) у шийку матки проникає лише 0,05-0,5 % (4—20 млн). Велика складчастість слизової оболонки шийки матки та густий в'язкий секрет затримують тут сперміїв. Істотну роль у їх просуванні по статевих органах самки відіграє моторика матки та яйцепроводів, яка різко зростає з настанням тічки та охоти. її підсилює виділюваний задньою долею гіпофіза окситоцин.
Крім того, у спермі окремих видів тварин, зокрема у секреті простати, є простагландини, що володіють утеротонічною дією. Нарешті, після еякуляції у спермі утворюється ацетилхолін, що підсилює моторику матки. Сильні ритмічні скорочення поздовжніх м'язів матки супроводжуються розслабленням циркулярних м'язів, що приводить до скорочення матки і розкриття цервікального каналу. Позитивно позначається на моториці матки присутність самця. Як установив М. Полянцев, перистальтичні та антиперистальтичні скорочення матки у корів, найкраще виражені під час статевого збудження, підсилюються при наближенні бугая. Моторику матки також підсилює попадання у піхву корів секретів додаткових статевих залоз бугая.
У дослідах на кобилах Г. В. Паршутін установив, що при підведенні жеребця до кобили у стані охоти її матка, що була до цього розслабленою, починає скорочуватися, спочатку скорочуються циркулярні м'язи, а через хвилину- поздовжні (циркулярні при цьому розслаблюються), внаслідок чого у порожнині матки наступає розрідження, що продовжується біля 3-6 хвилин. Завдяки такій засмоктуючій дії сперма досягає верхівки рогів матки протягом декількох секунд. При наступному розслабленні поздовжніх та скороченні циркулярних м'язів основна частина еякуляту засмоктується ще глибше.
Добре виражена засмоктуюча дія матки у свиней. Скорочення матки у них стимулюється окситоцином, що виділяється рефлекторно з гіпофіза у відповідь на подразнення рецепторів геніталій під час статевого акту чи штучного осіменіння. Якщо під час осіменіння корови або зразу після нього її ударити чи крикнути на неї, то адреналін, який при цьому виділяється, гальмує дію окситоцину, знижує скорочення матки і затримує просування сперміїв по статевих шляхах.
У 60-х роках минулого сторіччя група англійських біологів довела, що у жуйних у відповідь на подразнення рецепторів статевих органів під час нормально проведеного : ::меніння внаслідок антиперистальтичних скорочень та засмоктуючої дії матки відбувається "механічне закидання сперміїв" з цервікального каналу до верхівок рогів лтки. Для цього потрібно всього 3-5 хвилин.
Отже, просування сперміїв по статевих шляхах самки відбувається за рахунок ре-
: таксису, динаміки статевого акту та скорочень матки. У яйцепроводі додаються ще
війчастого епітелію та зростання тиску черевних стінок. З цього можна зробити
сновок, що швидкість переміщення сперміїв по статевих шляхах самки залежить
і а виду осіменіння, рухливості сперміїв, реакції самки на присутність самця, подраз-
і рецепторів геніталій під час статевого акту чи штучного осіменіння.
189
Розділ Є
Швидкість переміщення сперміїв з місця введення до яйцепроводів у різних ссавців складає від 15 сек. до 1-3 годин. Є дані, що у корови вони досягають яйцепроводів через 2,5 хв. після осіменіння, тоді як В. Нагаєв виявляв їх лише через 3-14 годин. У овець одні дослідники виявляли їх через 0,5-1 годину, інші - через 6 хв. після осіменіння. У кобил спермії з'являються у яйцепроводі через 30-60 хв., а у свиней при природному осіменінні спермії досягали яйцепроводів через 15 хв., проникали у них через 0,5-1 годину, а при штучному осіменінні - досягали яйцепроводів через 45 хв.. виявлялися у них через 1,5-2 години після осіменіння. У собак виявляли сперміїв у яйцепроводі через 25 сек. від початку еякуляції.
Проте у яйцепроводи попадає лише частина сперміїв, більшість їх затримується у складках шийки матки. Частина сперміїв попадає у вивідні протоки маткових залоз, інші аглютинуються. Нарешті, вхід у яйцепровід розкривається лише під кінець охоти.
На швидкість проходження сперміями до яйцепроводу впливає стан статевих шляхів та метод осіменіння.
Яйцепровід, як і уся статева система самки, перебуває під нейрогуморальним контролем, парадоксальність якого полягає у тому, що у передовуляційну фазу спермії рухаються у напрямку ампули яйцепроводів, а після овуляції - у протилежному напрямку. Найкращим часом для досягнення ампул є період охоти. Спермії, що досягай перешийка яйцепроводу, скупчуються тут у передовуляційну фазу. Є дані, що вони можуть навіть впадати тут у стан анабіозу і перебувати в ньому до 18 годин. Після овуляції під впливом фолікулярної рідини їх рухливість поновлюється. Згідно Т. Бішопа і А. Грея, при температурі 37 °С спермій бугая здійснює 9 ударів за сек., просуваючись при кожному ударі на 8,3 мкм, а за одну хв. - на 4 482 мкм.
Рух сперміїв по яйцепроводах забезпечується головним чином за рахунок рухів війчастого епітелію яйцепроводів та власних рухів сперміїв.
Після овуляції погіршуються умови для просування сперміїв до верхівок яйцепроводів, цервікальний канал звужується, стає звивистим, скорочення матки стають перистальтичними.
Згідно Р. Мартиненка, у свиней антиперистальтичні скорочення матки поступово наростають і досягають найбільшої інтенсивності через 24 години від початку охоти.
Виживання гамет у різних ділянках статевих органів. Результативність осіменіння буває тим вищою, чим менше часу пройде між уведенням сперми у статеві органи самки і овуляцією, яка у окремих тварин не співпадає з охотою, коли проводиться осіменіння. Більше того, у великої рогатої худоби овуляція відбувається через 15 годин після закінчення охоти. Ігнорування цими особливостями статевого циклу приводить до того, що нерідко осіменіння проводять передчасно чи запізно, спермії попадають у яйцепроводи або задовго до овуляції, або після неї, коли яйцеклітина або ще не може, або вже не може з'єднатися зі спермієм. Тому важливо знати, з одного боку, особливості виживання сперміїв у різних ділянках статевого апарату по періодах статевого циклу, а з іншого, оптимальний час осіменіння кожного виду тварин. Наявні дані наукових досліджень показують, що найгірші умови для виживання
190
Запліднення
сперміїв у піхві, де вони зберігають рухливість 2-6 годин (у кобил 4—4,5, корів 1-6). Це зв'язане з низьким рН середовища, активним фагоцитозом, наявністю тут мікроорганізмів, домішок клітин крові. При виникненні запальних процесів у піхві виживання тут сперміїв значно знижується, наприклад, при вагініті у корів спермії гинуть протягом першої години, у кіз - через 1 годину 45 хв. після введення.
Дані про виживання сперміїв у матці досить розбіжні. Одні вважають його досить високим, інші, навпаки, низьким, пояснюючи це наявністю тут, особливо у тварин з матковим типом осіменіння, лужного секрету, який активує рухливість сперміїв і вкорочує їх виживання. Хоча є дані, що виживання сперміїв у матці свиней та кобил, особливо у їх верхівках, сягає до 30-48 годин.
У яйцепроводах виживання сперміїв знижується до 6-9 годин.
Цікаві дані отримано у дослідах на кролях. Як відомо, овуляція у кролиць наступає приблизно через 10 годин після коїтусу. При штучному осіменіння кролиць високоякісною спермою з подальшим їх спаровуванням з вазектомованим самцем через 8 годин вагітність наступала у 90,9 % випадків. Якщо коїтус з вазектомованим самцем проводили через 20 годин після штучного осіменіння, то запліднення наступало лише у 12,9 % випадків. Нарешті, при коїтусі через 30 годин після осіменіння вагітність не наступала. Тобто, виживання сперміїв у статевих органах кролиці зберігається до 40 годин.
Найвище виживання сперміїв у шийці матки, особливо у самок з піхвовим ти-вом природного осіменіння. Тут спермії зберігають запліднюючу здатність найдовше, наприклад, у корови - до 30 годин, вівці - до 48 годин, хоча основна маса їх проходить через цервікальний канал досить швидко (15-50 хв). Тривалому зберіганню тут сперміїв сприяє велика складчастість слизової оболонки,та наявність густого слизу із слабо кислою реакцією. Дослідження цервікального слизу показали, що він складаться з глікопротеїду та плазми. Міцели глюкопротеїду розміщені паралельно і між ними можуть вільно просуватися спермії з прямолінійно-поступальним рухом. Та-і шу розташуванню міцел глікопротеїду сприяє поява у слизі високих концентрацій естрогенів (і низьких - прогестерону) під час тічки та охоти. У лютеїнову фазу слиз лає в'язким, розщепленим, що значно перешкоджає руху сперміїв. Завдяки цьому, а іапш під впливом секретів додаткових статевих залоз під час охоти, активні спермії ■■ганяються тут від мертвих та мікроорганізмів. Якщо ж осіменіння проводять після чення охоти, то спермії гинуть тут уже через 2-4 години, не досягаючи рогів шли. Отже, на всьому шляху від місця введення сперміїв до місця запліднення від-'- зветься "відсів" слабших сперміїв (елімінація). Першим "фільтром" для сперміїв є лгрвікальний слиз. Далі відбувається відсів сперміїв з ферментними дефектами.
Спермії, що проникли у матку, проштовхуються антиперистальтичними скорочен-
:: її до верхівок яйцепроводів. Тривалість антиперистальтичних скорочень матки
"—5 ГОДИН.
Яйцепровід розкривається під час овуляції і пропускає у корів та телиць 1 000 . -есміїв, у овець - 500. У яйцепроводах рух сперміїв активізується фертилізином фо-купярної рідини, що забезпечує синхронність входження у зону запліднення яйця і :нггмія.
191
Розділ 6
Виживання сперміїв у статевих органах самки, збереження ними тут запліднюючої здатності визначаються фізіологічним станом самки, який у свою чергу піддається змінам у зв'язку з умовами утримання та годівлі тварин. Так, якщо осіменіння тварин проводиться не під час охоти, то вже через декілька хвилин спермії піддаються аглютинації і гинуть. В. Парпгутін установив, що у статевих органах кобил з низькою вгодованістю спермії гинуть вже через 12-24 години, тоді як у кобил з середньою та доброю вгодованістю вони виживали до 48 годин. Окремі токсини, лікарські та інші речовини можуть проникати з кров'яного русла у статеві органи і виділювані ними секрети і позначатися на виживанні та запліднюючій здатності сперміїв. У дослідах з введенням лабораторним самкам рег он свинцю, нікотину, фосфору відмічено значне скорочення виживання сперміїв, у 50 % випадків виникали аборти. Знижували також виживання сперміїв введення самкам миш'яку, ртуті, колоїдального срібла та заліза.
Таким чином, виживання сперміїв у статевих органах самки визначається багатьма факторами, на першому місці серед яких слід поставити фізіологічний стан статевих органів та загальний стан здоров'я самки; зміни фізіологічного стану статевих органів, виникнення хворобливого стану чи, навпаки, одужання; неповноцінна чи, навпаки, повноцінна годівля викликають відповідні зміни у окремих ланках обміну речовин, характері і якості статевих секретів, тічкового слизу, що в свою чергу погіршує чи поліпшує виживання тут сперміїв. Безумовно, воно у свою чергу залежить від якості самої сперми, яка у прямій мірі залежить від фізіологічного стану плідника. стану його здоров'я, умов утримання, характеру годівлі та експлуатації.
У птахів спермії скупчуються у складках та криптах лійки яйцепроводу і зберігають запліднюючу здатність протягом 2-3-х тижнів. У окремих рептилій та риб їх виявляли живими у спеціальних сперміотеках протягом декількох місяців і навіть років.
Проте ефективність осіменіння залежить не лише від сперміїв, а й від виживання та біологічної активності яйцеклітини. А ці риси, на жаль, зберігаються у яйцеклітини значно коротше, ніж у сперміїв. Це, по-перше, зв'язане з тим, що яйцеклітина виходить з фолікула не повністю дозрілою, а у стані овоцита II порядку.
Другий поділ дозрівання яйцеклітини відбувається зразу після проникнення у неї спермія як наслідок активування яйця, що супроводжується двома морфологічними перетвореннями - відновленням мейозу та кортикальною реакцією. Якщо ж запліднення не наступило, то яйцеклітина швидко старіє і гине. По-друге, прозора оболонка яйцеклітини, на відміну від оболонки спермія, не проникна для поживних речовин, її енергетичні запаси досить обмежені. Згідно А. Лопиріна, старіння яйця супроводжується відповідними біохімічними змінами, які несприятливо позначаються на наступному онтогенезі. Тому при визначенні тривалості життя яйцеклітини слід враховувати не лише період, протягом якого можливе її запліднення (він складає для яйцеклітини близько 12-ти годин), але й час, протягом якого зберігається потенціальна здатність яйця до подальшого нормального онтогенезу, який, мабуть, не перевищує 5 годин.
Під час овуляції яйцеклітина, разом з клітинами променевого епітелію (кумулюс) виходить у простір, оточений пристінковою очеревиною таза та широкою черевною зв'язкою, у якій відкривається своїм широким кінцем яйцепровід.
192
Запліднення
У багатьох тварин кайма яйцепроводу тісно зв'язана з яєчником; внаслідок наповнення її судин кров'ю під час охоти вона стає щільною і притягується до яєчника, і навіть охоплює його. Згідно О. Квасницького, на місці розриву фолікула виявляється прозора драглиста маса, яка легко виймається з фолікула у вигляді тонкого ніжного тяжа, у середині якого знаходиться яйцеклітина, оточена променевим вінцем. Війчастими рухами клітин яйцепроводу цей тяж втягується у його просвіт. У тих же тварин, де кайма слабо розвинена (корова, вівця, коза), проникненню яйцеклітини у яйцепровід сприяють потоки рідини з черевної порожнини у матку. Як установив В. Груздєв, ці потоки скеровуються у напрямку матки коливальними рухами війчастого епітелію. Як тільки овоцит входить у яйцепровід, війковий епітелій, що раніше коливався у напрямку ампули яйцепроводу, тепер починає рухатися у зворотному напрямку, тобто від яєчника до матки. Переміщенню з потоками рідини яйцеклітини у яйцепровід сприяє також підвищення внутрішньочеревного тиску під час овуляції К. Скробанський).
Проте, це не виключає можливості деякої міграції яйцеклітини у черевній порожнині. У жінок, наприклад, у більшості випадків трубної вагітності овуляція відбувається у правому яєчнику, а плід розвивається у лівій трубі, що пояснюють зовнішньою черевною) чи внутрішньою (матково-трубною) міграцією яйцеклітини. Отже, після овуляції яйцеклітина може деякий час плавати у капілярних щілинах між петлями кишечника, або ж переміщатися через тіло матки з одного яйцепроводу в інший.
Істотний вплив на моторику яйцепроводів мають стероїдні гормони, катехоламі-ни. окситоцин та інші речовини. Яйцеклітина переміщується по ампулі яйцепроводу рухами епітелію, а у ділянці перешийка - також за рахунок скорочень його мускульного шару. До овуляції тут переважають антиперистальтичні, а після овуляції - перистальтичні скорочення.
Керуючись даними з фізіології овуляції, у тому числі множинної, можна зробити зисновок, що просування яйцеклітин по яйцепроводах триває від однієї до трьох діб, проте здатність до запліднення вони зберігають у межах 4-6 годин.
Проникнення спермія у яйцеклітину та розвиток зиготи. Яйцеклітина, що вийшла з фолікула, кулястої форми, у ній розрізняють ядро та цитоплазму (ооплазму), .уточені жовтковою оболонкою (оолемою) та прозорою оболонкою. Ззовні вона оточена клітинами променевого вінця, з'єднаними між собою в'язкими драглями, основу яких складає гіалуронова кислота (рис. 39). Для запліднення яйцеклітина повинна звільнитися від навколишніх клітин. У 1931 р. Ямане виявив у сперміях фермент гіалуронідазу, що розчиняє гіалуронові драглі. Вважають, що місія гіалуронідази І: литься до деполімеризації гіалуронової кислоти, руйнування міжклітинного ма-ттиксу, який з'єднує клітини променевого вінця, щоб дозволити окремому спермію ~: вити між фолікулярними клітинами до яйця. Згідно Остіна та Бішопа, для проникнення спермія у яйцеклітину не потрібно повного оголення її від клітин променевого інця, а лише виникнення між ними щілини.
193
Розділ
Виживання сперміїв у статевих органах самки, збереження ними тут заплідни ■ ючої здатності визначаються фізіологічним станом самки, який у свою чергу піддається змінам у зв'язку з умовами утримання та годівлі тварин. Так, якщо осіменін:-:: тварин проводиться не під час охоти, то вже через декілька хвилин спермії піддають с • аглютинації і гинуть. В. Паршутін установив, що у статевих органах кобил з низькок вгодованістю спермії гинуть вже через 12-24 години, тоді як у кобил з середньою та доброю вгодованістю вони виживали до 48 годин. Окремі токсини, лікарські та інс: речовини можуть проникати з кров'яного русла у статеві органи і виділювані ним секрети і позначатися на виживанні та запліднюючій здатності сперміїв. У дослідах : введенням лабораторним самкамрег оз свинцю, нікотину, фосфору відмічено значн: скорочення виживання сперміїв, у 50 % випадків виникали аборти. Знижували тако>і виживання сперміїв введення самкам миш'яку, ртуті, колоїдального срібла та заліза.
Таким чином, виживання сперміїв у статевих органах самки визначається багатьма факторами, на першому місці серед яких слід поставити фізіологічний стан статевих органів та загальний стан здоров'я самки; зміни фізіологічного стану статевих органів, виникнення хворобливого стану чи, навпаки, одужання; неповноцінна чи навпаки, повноцінна годівля викликають відповідні зміни у окремих ланках обмін;. речовин, характері і якості статевих секретів, тічкового слизу, що в свою чергу погіршує чи поліпшує виживання тут сперміїв. Безумовно, воно у свою чергу залежнії від якості самої сперми, яка у прямій мірі залежить від фізіологічного стану плідник; стану його здоров'я, умов утримання, характеру годівлі та експлуатації.
У птахів спермії скупчуються у складках та криптах лійки яйцепроводу і зберігають запліднюючу здатність протягом 2-3-х тижнів. У окремих рептилій та риб їх виявляли живими у спеціальних сперміотеках протягом декількох місяців і навіть років
Проте ефективність осіменіння залежить не лише від сперміїв, а й від виживанні та біологічної активності яйцеклітини. А ці риси, на жаль, зберігаються у яйцеклітини значно коротше, ніж у сперміїв. Це, по-перше, зв'язане з тим, що яйцеклітин, виходить з фолікула не повністю дозрілою, а у стані овоцита II порядку.
Другий поділ дозрівання яйцеклітини відбувається зразу після проникнення у не: спермія як наслідок активування яйця, що супроводжується двома морфологічними перетвореннями - відновленням мейозу та кортикальною реакцією. Якщо ж запліднення не наступило, то яйцеклітина швидко старіє і гине. По-друге, прозора оболонка яйцеклітини, на відміну від оболонки спермія, не проникна для поживних речовин, її енергетичні запаси досить обмежені. Згідно А. Лопиріна, старіння яйця супроводжується відповідними біохімічними змінами, які несприятливо позначаються на наступному онтогенезі. Тому при визначенні тривалості життя яйцеклітини слід враховувати не лише період, протягом якого можливе її запліднення (він складає для яйцеклітини близько 12-ти годин), але й час, протягом якого зберігається потенціальна здатність яйця до подальшого нормального онтогенезу, який, мабуть, не перевищує 5 годин.
Під час овуляції яйцеклітина, разом з клітинами променевого епітелію (кумулюс виходить у простір, оточений пристінковою очеревиною таза та широкою черевнок зв'язкою, у якій відкривається своїм широким кінцем яйцепровід.
192
Запліднення
У багатьох тварин кайма яйцепроводу тісно зв'язана з яєчником; внаслідок наповнення її судин кров'ю під час охоти вона стає щільною і притягується до яєчника, і навіть охоплює його. Згідно О. Квасницького, на місці розриву фолікула виявляється прозора драглиста маса, яка легко виймається з фолікула у вигляді тонкого ніжного тяжа, у середині якого знаходиться яйцеклітина, оточена променевим вінцем. Війчастими рухами клітин яйцепроводу цей тяж втягується у його просвіт. У тих же тварин, де кайма слабо розвинена (корова, вівця, коза), проникненню яйцеклітини у яйцепровід сприяють потоки рідини з черевної порожнини у матку. Як установив В. Груздєв, ці потоки скеровуються у напрямку матки коливальними рухами війчастого епітелію. Як тільки овоцит входить у яйцепровід, війковий епітелій, що раніше коливався у напрямку ампули яйцепроводу, тепер починає рухатися у зворотному напрямку, тобто від яєчника до матки. Переміщенню з потоками рідини яйцеклітини у яйцепровід сприяє також підвищення внутрішньочеревного тиску під час овуляції К. Скробанський).
Проте, це не виключає можливості деякої міграції яйцеклітини у черевній порожнині. У жінок, наприклад, у більшості випадків трубної вагітності овуляція відбуваються у правому яєчнику, а плід розвивається у лівій трубі, що пояснюють зовнішньою черевною) чи внутрішньою (матково-трубною) міграцією яйцеклітини. Отже, після шуияції яйцеклітина може деякий час плавати у капілярних щілинах між петлями кишечника, або ж переміщатися через тіло матки з одного яйцепроводу в інший.
Істотний вплив на моторику яйцепроводів мають стероїдні гормони, катехоламі-:-:;:. окситоцин та інші речовини. Яйцеклітина переміщується по ампулі яйцепроводу т ухами епітелію, а у ділянці перешийка - також за рахунок скорочень його мускульного шару. До овуляції тут переважають антиперистальтичні, а після овуляції - перистальтичні скорочення.
Керуючись даними з фізіології овуляції, у тому числі множинної, можна зробити і исновок, що просування яйцеклітин по яйцепроводах триває від однієї до трьох діб, щ оте здатність до запліднення вони зберігають у межах 4-6 годин.
Проникнення спермія у яйцеклітину та розвиток зиготи. Яйцеклітина, що ви-Ьшга з фолікула, кулястої форми, у ній розрізняють ядро та цитоплазму (ооплазму), гочені жовтковою оболонкою (оолемою) та прозорою оболонкою. Ззовні вона ото-сяа клітинами променевого вінця, з'єднаними між собою в'язкими драглями, основу яких складає гіалуронова кислота (рис. 39). Для запліднення яйцеклітина пови-в звільнитися від навколишніх клітин. У 1931 р. Ямане виявив у сперміях фермент іиуронідазу, що розчиняє гіалуронові драглі. Вважають, що місія гіалуронідази і миться до деполімеризації гіалуронової кислоти, руйнування міжклітинного ма-::-;%, який з'єднує клітини променевого вінця, щоб дозволити окремому спермію йти між фолікулярними клітинами до яйця. Згідно Остіна та Бішопа, для проник-евня спермія у яйцеклітину не потрібно повного оголення її від клітин променевого ї.ніія. а лише виникнення між ними щілини.
193
Розділ б
Повний розпад кумулюс; відбувається лише через де який час після проникнення спермія у яйцеклітину, вна слідок збільшення кількості сперміїв (і концентрації гіа- луронідази) у зоні находжен ня яйцеклітини, аутолітичних процесів та дії йонів бікарбо нату у секретах фаллопієвих труб. Звільнення зиготи від фолікулярних клітин має важ ливе значення для нормально го обміну газів та метаболітів. Рис. 39. Яйцеклітина. Виходячи з цих міркувань.
І. Соколовська запропонувала
вважати звільнення яйцеклітини від променевого вінця (денудація) першою стадією запліднення, хоча більшість учених вважають цей феномен підготовчою стадією.
Сам процес запліднення включає такі стадії: 1) проникнення спермія крізь прозору оболонку; 2) активування яйця і блокада поліспермії; 3) утворення чоловічого та жіночого пронуклеусів: 4) заміна пронуклеусів хромосомними групами; 5) об'єднання двох хромосомних груп (сингамія).
(zona pellucida)
Механізм проникнення спермія через прозору оболонку повністю не з'ясований. Одні автори вважають, що у ньому беруть участь багато сперміїв, частина з яких навіть можуть занурюватися голівками у прозору оболонку, інші ж затримуються на її поверхні. Прихильники протилежної точки зору вважають, що як тільки спермій пройде між фолікулярними клітинами і доторкнеться голівкою прозорої оболонки чи може навіть зіллється з нею боковою поверхнею голівки, під впливом виділюваного ним ферменту зоналізину у прозорій оболонці виникає щілина, крізь яку він проникає у яйцеклітину. Прозора оболонка (гопареііисісіа) - це густе плетиво глікопротеїдів, у петлях якого є вуглеводні компоненти- кортикальні гранули 2Р1, 2Р2 і 2РЗ, з яких останній (2РЗ) є специфічним рецептором для спермія. Прикріпившись до нього, спермій виділяє фермент акрозин (акросомна реакція), під впливом якого у прозорій оболонці виникає канал для проникнення спермія. Далі вступає у дію другий глікопротеїд (2Р2). що викликає затвердіння прозорої оболонки і захищає яйцеклітину від проникнення інших сперміїв. Наступає "блок поліспермії". Під час проникання спермія у глибину цитоплазми яйця завершується другий поділ мейозу та дозрівання яйцеклітини, виділяється друге напрямне тільце (рис. 40).
Момент доторкання голівки спермія до яйця є початковою ланкою у ланцюгу послідовних змін, що розпочинаються з активування яйцеклітини: у неї зростає проник -
194
Запліднення
ність прозорої оболонки для йонів К* та Са++; активується синтез ліпідів, білків, змінюється в'язкість та інші колоїдальні властивості білків; знижується мембранний потенціал яйця; жовткова оболонка потовщується, відтісняється від поверхні ооплазми, утворюючи перивітеліновий простір.
У деяких видів тварин (хом'як, сука, вівця) реакція прозорої оболонки наступає дуже швидко, тоді як у інших (наприклад, у щурів) вона сповільнена. У кролів не відмічено реакції прозорої оболонки, тому у пе-гивітеліновому просторі у них може . куштуватися багато сперміїв. Описані випадки проникнення у яйцеклітину кролиці до 200 "зайвих" сперміїв.
Перивітеліновий простір віднос-
: малий, тому голівка спермія, що
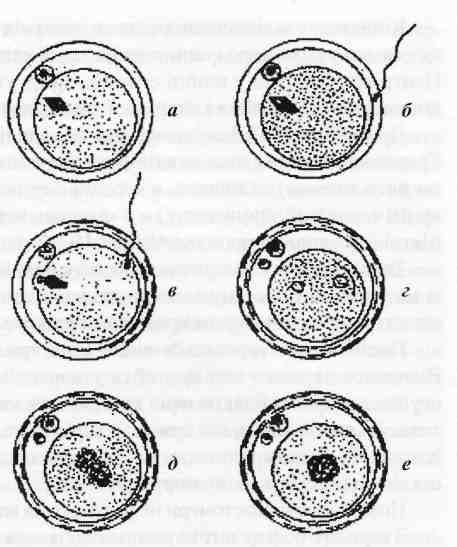
Рис. 40. Стадії запліднення:
а - незапліднена яйцеклітина, б - проникнення спермія через прозору оболонку, в - активування овоцита, г - формування чоловічого та жіночого пронуклеусів, д - заміна пронуклеусів хромосомними групами, є - сингамія.
проник сюди, торкається жовткової
г міонки і наче прилипає до неї.
Після деякої паузи спермій проникає
: :зь жовткову оболонку у цитоплаз-
<у. Вважають, що цей процес нага-
цує фагоцитоз або захоплення аме-
: часток їжі. Він суворо специфічний, у ньому може брати участь лише спермій .:: го виду.
Відбуваються зміни голівки спермія, вона з лопатоподібної стає циліндричною.
Велику роль у цих змінах спермія і яйця відіграє секрет яйцепроводу, наявні у
і : му інгібітори ферментів акрозину та трипсину, рівень яких зростає перед овуля-
_: по, тоді значно знижується під час овуляції і знову зростає у наступні 1-2 дні. Ці
ібітори ферментів сприяють настанню у певний час овуляції і забезпечують захист
: ерментів сперміїв, що розпадаються.
Спермії, що зосередилися у верхній третині яйцепроводу, перебувають у стані івабіозу і лише за декілька годин до овуляції піддаються капацитації
Зразу після проникнення спермія крізь жовткову оболонку його голівка відокремлюся від джгутика, збільшується в об'ємі (приблизно у 20 разів) і рухається у напрямку
?. яйцеклітини. Воно трансформується у жіночий пронуклеус, а ядро (голівка) спер-'-•:л - у чоловічий пронуклеус. Чоловічі та жіночі пронуклеуси зближуються, зменшуються у об'ємі і втрачають свої оболонки, перетворюючись у хромосомні набори.
Розділ Є
Кінцевою стадією запліднення є сингамія - об'єднання хромосомних наборів самця і самки у одне ядро і виникнення нової клітини - зиготи з геномом нового індивіда. Центріолі, що були у шийці спермія, зразу утворюють веретено, у якому розташовуються хромосоми яйця і спермія, і зигота виявляється готовою до початку поділу.
Thibault
Дослідження ТкіЬаиІі показали, що запліднення - це не одномоментний процес. Тривалість перших двох стадій (проникнення спермія крізь прозору оболонку та зміни його голівки) становить в середньому по 3 години, третьої (перетворення голівки на чоловічий пронуклеус) - 4-5 годин, четвертої (зближення пронуклеусів) - 8-9, п'ятої (об'єднання пронуклеусів) - 10-12 годин.
Таким чином, запліднення виконує подвійну роль: по-перше, воно виводить яйце із загальмованого стану, а, по-друге, воно приводить до утворення диплоїдної клітини зиготи з повним набором хромосом батьківської та материнської спадковості.
Поділ зиготи торкається лише ядра при збереженні цілості прозорої оболонки. Внаслідок першого поділу зиготи утворений зародок має два бластомери. Кожне наступне покоління бластомерів виявляється меншим попереднього, тому цей поділ називають дробленням, що триває у корів та телиць близько 8-ми діб. Приблизно через кожних 24 години кількість бластомерів подвоюється, хоча у ссавців уже на ранніх стадіях ця чіткість може порушуватися.
Новоутворені бластомери не розходяться на боки, а залишаються компактною масою.
З першого поділу зиготи розпочинається складний процес внутрішньоутробного розвитку нового організму, що завершується врешті формуванням життєздатного плода.
Таким чином, у процесі запліднення беруть участь, з одного боку, - мільйони сперміїв, а з другого - лише одна яйцеклітина. І доля майбутнього зародка залежить, у першу чергу від якості яйцеклітини. Серед мільйонів сперміїв завжди знайдеться якась кількість високоякісних, зате, якщо яйцеклітина виявиться неповноцінною, то запліднення або ж не наступить, або новоутворений зародок буде нежиттєвим.
Для запліднення у кінцевому рахунку потрібен один спермій. Це явище називають моноспермією. Проте у науці відомі випадки поліспермії. Так, у цитоплазму яйця голуба при заплідненні проникає 15-25 сперміїв, у курей - 5-6. Це ж саме стосується риб. Подібна поліспермія вважається нормальною. Проникнення ж зайвих сперміїв у яйце ссавців розцінюється патологічною поліспермією.
Проникнення зайвих сперміїв у яйця тварин, яким властиве моноспермне запліднення, викликає порушення у їх розвитку. При нормальному заплідненні зливається два пронуклеуси і поновлюється диплоїдний набір хромосом, а при поліспермії виникає декілька пронуклеусів, але у злитті ядер (каріогамії) може брати участь або лише дві групи хромосом, або гомологічні хромосоми усіх пронуклеусів, або ж зайві пронуклеуси просто елімінуються і запліднення завершується нормально. Якщо ж виникає справжня поліспермія, то внесені разом з іншими сперміями центросоми утворюють додаткові центри дроблення, і яйце дробиться потворно зразу на декілька бластомерів.
Однією з причин поліспермії є старіння яйцеклітини. У міру її старіння у неї все легше може проникнути зайва кількість сперміїв. Патологічна поліспермія приводить звичайно до ранньої ембріональної смертності чи вироджень.
196
Запліднення
Моноспермність запліднення у ссавців регулюється зменшенням кількості сперміїв, що можуть дійти до місця запліднення, і специфічною реакцією прозорої оболонки яйцеклітини.
Досі не відомо, чи здатні перші спермії, що досягли яйцеклітини, запліднити її. Для того, щоб взяти участь у заплідненні яйцеклітини, у сперміях повинні наступити відповідні морфологічні зміни, що забезпечують набуття ними здатності проникати у яйцеклітину. Ці зміни сперміїв було виявлено у 50-х роках XX ст. на різних видах тварин - Нойес - на щурах (1953), Брейден і Остін - на мишах (1954), Ченг і Шиф-іер - на хом'яках (1957), Меттнер - на вівцях (1963). У дослідах іп уіїго було помічено, що спермії проникали у яйце лише після додавання у пробірку обрізків тканин шцепроводу, матки чи слизової оболонки яйцепроводу, або ж коли у проби додавали спермії, які хоча би 8 годин перебували у рогах матки чи яйцепроводах. Ченг і Остін прийшли до висновку, що спермії набували запліднюючої здатності лише після 8-ми годин перебування у секреті матки та яйцепроводу. За цей час у них відбуваються певні зміни, які отримали назву капсщитацїі. Суть цих змін полягає у акросомній реакції, тобто у набуханні передньої частини акросоми; злитті її зовнішньої мембрани з плазматичною мембраною наступною вакуолізацією; скиданні акросоми; звільненні сперміїв від деяких макромолекул та обволікаючих антигенів, що перешкоджають ви-стленню ензимів; вивільненні вмістимого акросоми.
Детермінація статі. Під час запліднення відбувається не тільки формування генотипу майбутнього індивіда, а й визначення його статі.
Як згадувалось вище, овогонії та сперматогонії містять властивий для свого виду ди-плоїдний (2п) набір хромосом, у тому числі дві статеві: XX - у самок і ХУ - у самців.
Під час редукційного поділу овоцита І порядку його диплоїдний хромосомний на-гір замінюється гаплоїдним, внаслідок чого виникає авоцит 2-го порядку, що містить, наприклад, у корів лише 30 хромосом, з них 29 аутосом і одна статева (X). Те ж саме стосується сперміїв, лише половина з них містить статеву хромосому X, а друга по-і: вина - хромосому У.
Стать майбутнього індивіда залежить від того, який вид спермія — носій хромосоми X чи У з'єднається з яйцеклітиною. У першому випадку хромосомний набір гїгготи включатиме статеві хромосоми XX, тому з неї згодом сформується самка, а другому випадку - хромосоми ХУ, що дасть початок розвитку самця. Тобто, стать
: ла визначається статевими хромосомами спермія.
Отже, генетично стать визначається під час запліднення, анатомічно - вона оформлять ся у великих тварин на 7-10-му тижні розвитку ембріона під впливом статевих гормонів і у плода формуються соматичні риси І, II та Ш-го порядку. Риси 1-го порядку - це готи. П-го - внутрішні і зовнішні статеві органи; риси Ш-го порядку — статеві рефлекси.
Проникнення спермія у яйцеклітину та утворення зиготи є першим етапом розви-~у нового організму. Проте виникнення багатьох організмів може відбуватися і без
т лнення, а шляхом партеногенезу. У бджіл, наприклад, самці розвиваються з яєць іез .часті сперміїв. У одній родині коловерток самців узагалі немає. Самка дафнії за :>:-: життя може виділити близько ста яєць, які розвиваються партеногенетично.
197
Розділ 6
Яйцеклітина має всі основні елементи, необхідні для початку розвитку.
При партеногенезі диплоїдний набір хромосом утворюється за рахунок зворотного втягування полярного тільця або ж випадіння редукційного поділу. Ще у XIX ст. А. Тихонову вдалося викликати штучний партеногенез у яєць шовковичного шовкопряда, подразнюючи їх слабкою кислотою, механічно, термічно.
Що спільного між названими прийомами і дією спермія на яйце?
Спермій, проникаючи крізь прозору оболонку, викликає у яйцеклітині якусь початкову реакцію, що швидко прогресує. У дослідах Берта, при обережному введенні мікропіпеткою живого спермія у цитоплазму яйця, запліднення не наступало.
М. Олсен довів можливість штучного партеногенезу у птахів. Виведено навіть породу індичок, у яких приблизно у половині випадків спостерігається природний партеногенез. X. Пінкусу вдалося добитися партеногенетичного розвитку яєць кролиці. П. Хегестрем виявив у яєчнику жінки яйця на перших стадіях дроблення і ранню бластулу, а В. Хватов виявив яйцеклітину на стадії 8-ми бластомерів у фолікулі 23-річної дівчини; осіменіння і запліднення у даному випадку були виключені.
Проблема партеногенезу особливо актуальна при клонуванні нащадків високопродуктивних тварин. Адже з величезної кількості наявних у яєчнику яйцеклітин лише одиниці використовуються для відтворення нащадків.
Близнюки (двійнята). Якщо під час овуляції у одноплідних тварин виходить декілька яйцеклітин, то у випадку їх запліднення і виношування народжуються різнояйце-ві близнята (двійнята), трійнята і т. д. Якщо ж овулює один фолікул і зигота, що виникає в результаті запліднення, ділиться зразу не на два бластомери, а на дві окремі частини, з кожної з яких розвивається окремий плід, то говорять про однояйцевих близнят.
(superfecundatio)
Якщо ж самці бувають однієї породи, то розрізнити приплід практично не можливо.