

19.2. Научение
19.2.1. ФОРМЫ НАУЧЕНИЯ
Существует несколько классификаций научения. Целесообразно все их объединить в четыре основные группы преимущественно по критерию активности животного или человека в ходе научения: а) пассивное (реактив? ное) научение, б) активное (оперантное) научение (operatio — действие), в) научение- с помощью наблюдения, г) инсайт.
А. Пассивное (реактивное) научение имеет место во всех случаях, когда организм пассивно {не прилагая целенаправленных усилий) реагирует на какие-то внешние факторы, в результате чего в нервной системе формируются новые следы памяти. Пассивным научением являются следующие формы.
1. Привыкание — угасание ориентировочной реакции (рефлекса «что такое?» по И.П.Павлову). Если раздражитель многократно повторяется и не имеет особого значения для организма, то последний прекращает на него реагировать, развивается привыкание (габи-туация). Ориентировочная реакция угасает. Например, появление звука в лаборатории, где находится подопытное животное (собака), первоначально вызывает у нее ориентировочную реакцию: поворот головы, ушей в сторону источника звука. После многократного повторения звука, не сопровождающегося другими воздействиями на животное, последнее прекращает на него реагировать. У человека и животных новый раздражитель вызывает не только соматические, но и веге-
тативные реакции: изменение частоты сердечных сокращений, десинхронизацию ЭЭГ, изменение частоты и глубины дыхания. Если оказывается, что раздражитель не имеет значения для организма, то при последующем его повторении угасают и вегетативные, и соматические реакции. Так, человек, живущий на шумной городской улице, не обращает внимания на посторонний шум транспорта ночью, несмотря на него, крепко спит.
Угасание ориентировочной реакции — это не только самая простая, но, вероятно, и самая распространенная форма научения у человека и животных. За счет него нам удается игнорировать раздражители, не несущие никакой новизны и не имеющие для нас значения, сосредоточивая внимание на более важных явлениях. Привыкание всегда специфично в отношении стимула: если человек не замечает уличного шума, он проснется либо от неожиданного звука на его фоне, либо от внезапно наступившей тишины. Привыкание — это не утомление, а особый приспособительный нервный процесс, который не следует путать с адаптацией анализаторов, т.е. снижением их чувствительности при непрерывной стимуляции.
-
Сенсибилизация — усиление реакции ор ганизма на повторяющийся стимул, если он вызывает каждый раз неприятные ощущения. Например, капли воды из крана, жужжание назойливой мухи или пронзительный писк комара, многократно повторяющийся, ста новятся непереносимыми, неприятными. В данном случае научение носит негативный характер и выражается в соответствующих поведенческих реакциях типа стимул—ответ (после нескольких повторений действия раз дражителя).
-
Импринтинг — запечатление в памяти новорожденного окружающей действитель ности. Импринтинг — особая форма ассоци ативного научения, основанная на врожден ной предрасположенности к фиксации опре деленных сочетаний раздражителей и возни кающих ответных реакций в ранний период развития организма.
-
Классические условные рефлексы, по И.П.Павлову — положительные и отрицатель ные (см. разделы 19.2.2; 19.2.3; 19.2.4; 19.2.6).
Б. Активное (оперантное) научение -— научение, в ходе которого организм добивается полезного результата с помощью активного поведения. Имеется три основных вида научения — метод проб и ошибок, инструментальный условный рефлекс и самораздражение.
1. Метод проб и ошибок. Американский ученый Э.Торндайк (1890), видный бихевио-
507
рист (англ. behaviour — поведение) помещал голодных кошек в так называемые проблемные клетки, которые открывались в том случае, если кошка предпринимала какие-то определенные действия: тянула за веревку, приподнимала запорный крючок. Когда кошка выходила из клетки, она получала пищу. По мере повторения процедуры выхода из клетки, иначе увеличения числа проб и ошибок, скорость выполнения задачи возрастала.
-
Инструментальный условный рефлекс — научение.действию с помощью вознагражде ния (подкрепления). Например, при помеще нии в клетку Скиннера, где находится светя щийся пластмассовый кружок, голубь полу чает зернышко; если он повернет голову в сторону кружка — он вновь получает зер нышко; если голубь случайно клюнет кру жок, он снова получает зернышко и т.д. Вскоре голубь научается ударять клювом по кружку за вознаграждение. Подобное науче ние может проводиться с целью избавления или избегания неприятного воздействия, на пример раздражения электрическим током, пропускаемым через металлическую сетку пола. Так, животное (крыса) по световому сигналу нажимает на рычаг и выключает электрический ток, чтобы избежать раздра жения. Крыса также научается реагировать на световой раздражитель: нажимает на рычаг, чтобы получить пищу. т.е. она исполь зует какой-то инструмент — отсюда и назва ние этого вида научения. Дети быстро учатся говорить, когда родители одобряют их при правильном произношении отдельных звуков и слов. В случае неправильного произноше ния слова дети не получают подобного под крепления, в результате чего эти слова посте пенно исчезают из употребления.
-
Самораздражение структур мозга для по лучения удовольствия — положительных эмоций.
В, Научение путем наблюдения. Различают два вида подобного научения.
-
Простое подражание. Например, обезь яна в общении с исследователями научилась мыть банан перед едой, не понимая зачем это делается.
-
Викарное научение также осуществляет ся посредством наблюдения, но при этом результат действия оценивается. Такое на учение свойственно только человеку. Осо бенно часто оно используется детьми, при чем в раннем онтогенезе оно является пре имущественно подражательным. С возрас том викарное научение начинает все больше превалировать над подражательным науче нием.
/. Научение путем инсайта (озарение) —
внезапное нестандартное правильное решение задачи: иногда после отдельных проб и ошибок возникает идея результативного действия, которое может совершенствоваться в процессе достижения цели. Этот вид научения является следствием объединения опыта, накопленного в памяти, с той информацией, которой располагает индивидуум при решении проблемы. Так. находящаяся в клетке обезьяна в эксперименте с недосягаемым для нее бананом сначала пытается достать плод одной палкой в виде трубки, потом другой такой же, Но обе палки оказываются слишком коротки. Тогда она прекращает свои попытки, разглядывает находящиеся возле нее предметы и вдруг быстро проделывает логическую совокупность действий: схватывает обе палки, вставляет их одну в другую, просовывает между прутьями клетки и достает банан.
Д. Экстраполяционные рефлексы [Кру-шинский Л.В., 1960] состоят в способности животного определять направления дальнейшего перемещения значимого для него раздражителя, например перемещение пищи, движущейся по прямой линии после исчезновения из поля зрения. Способность к экстраполяции хорошо развита у птиц, хищных млекопитающих, дельфинов.
Следует подчеркнуть, что в конкретных ситуациях для достижения того или иного полезного приспособительного результата индивидуум чаще всего реализует не один, а несколько видов научения. Частное научение (обучение игре на музыкальном инструменте, обучение работе на компьютере и др.) по своей структуре всегда является комплексным.
19.2.2. ОСНОВНЫЕ ЗАКОНОМЕРНОСТИ ч ОБРАЗОВАНИЯ И ПРОЯВЛЕНИЯ УСЛОВНЫХ РЕФЛЕКСОВ И ИХ ХАРАКТЕРИСТИКА. МЕТОДЫ ИССЛЕДОВАНИЯ ВНД
А. Выработка условных рефлексов.
-
Необходимо неоднократное совпадение во времени действия индифферентного раз дражителя с безусловным.
-
Условный стимул должен предшество вать безусловному.
-
Условный рефлекс образуется на базе безусловного (врожденного) рефлекса.
Образованию условных рефлексов способствуют следующие факторы (условия).
• Оптимальное состояние организма (здоровье), в первую очередь состояние коры большого мозга.
508
-
Функциональное состояние нервного центра безусловного рефлекса (для пище вых рефлексов — наличие пищевой по требности).
-
Отсутствие посторонних сигналов как из внешней среды, так и от внутренних орга нов (кишечник, мочевой пузырь и др.).
-
Оптимальное соотношение силы условно го и безусловного раздражителей. Для об разования и сохранения условных рефлек сов существует определенный диапазон величин подкрепления — минимальный (пороговый), оптимальный, максималь ный.
В процессе выработки условного рефлекса выявлены периоды его образования:
-
скрытый период, когда условные ответы не возникают, несмотря на неоднократ ные сочетания условного и безусловного раздражителей;
-
период неустойчивых условных рефлек сов, когда не все условные раздражители сопровождаются появлением условных от ветов;
-
период генерализации, когда условные ре акции возникают на все посторонние сиг налы, помимо того, на который вырабаты вается рефлекс;
-
период специализации, когда условный ответ возникает только на тот условный сигнал, на который шла выработка;
-
финальная стадия — появление условного рефлекса относительно стабильной вели чины.
Б. Компоненты условного рефлекса. Любой рефлекс представляет собой сложно организованный поведенческий акт. Так, раздражение рецепторов пищевого безусловного рефлекса сопровождается рядом эффекторных ответов (компонентов) — дыхательным, двигательным, секреторным и др. Поскольку условный рефлекс образуется на базе безусловного и его воспроизводит, то и он состоит из ряда компонентов. На начальных этапах образования условного рефлекса в период изолированного действия условного сигнала отмечается определенная очередность появления компонентов: вначале обнаруживается сопряженный компонент (дыхательный, сосудистый, сердечный и др.), имеющий значение для энергетического обеспечения последующей специфической деятельности, затем появляется специфический поведенческий (соматический) компонент, состоящий из позно-тонических и фазных реакций, обеспечивающих приближение животного к пи-
щевой подкормке; позже в целостную услов-норефлекторную деятельность дополнительно включаются иные компоненты, например секреторные и моторные проявления желудочно-кишечного тракта.
В. Сходство и различия условных и безусловных рефлексов. Сходство этих рефлексов состоит в том, что их дуги имеют афферентную, центральную и эфферентную части; оба типа рефлексов в биологическом отношении имеют приспособительное значение. Имеется ряд отличий условных рефлексов от безусловных.
-
Безусловные рефлексы — врожденные, ус ловные — приобретаются в течение жизни, поэтому первые отражают видовые особенности организма, а вторые — инди видуальные.
-
Безусловные рефлексы относительно по стоянны в течение жизни особи, услов ные — образуются, изменяются и исчеза ют, когда они становятся неадекватными условиям жизни особи.
-
Безусловные рефлексы реализуются по анатомическим путям, определенным ге нетически, а условные —■ по функцио нально организующимся временным (за- мыкательным) связям.
-
Безусловные рефлексы свойственны всем уровням центральной нервной системы и преимущественно осуществляются ее низ шими отделами (спинной мозг, стволовый отдел, подкорковые ядра). Условные реф лексы для своего образования и реализа ции требуют целостности коры больших полушарий, особенно у высших млекопи тающих.
-
Каждый безусловный рефлекс имеет свое специфическое рецептивное поле и спе цифические раздражители. Условные реф лексы могут образовываться с любого ре цептивного поля на самые разнообразные раздражители.
-
Безусловные рефлексы возникают на дей ствие наличного раздражителя, которого уже нельзя избежать, а условные — при спосабливают организм к действию сти мула, которое еще предстоит испытать, т.е. имеют сигнальное (предупредитель ное) значение.
Г. Методы исследования ВНД. Для изучения закономерностей ВНД, особенностей формирования и протекания условных рефлексов чаще других используются следующие методы.
1. Методы разрушения и раздражения отдельных структур мозга, нацеленные на вы-
509
яснение основы условнорефлекторной деятельности. В последние годы стали использоваться методы холодового выключения структур головного мозга. Этот метод позволяет визуализировать пространственно-временную мозаику процессов мозга при образовании условного рефлекса в разных функциональных состояниях.
-
Фармакологические методы, связанные с введением различных биологически актив ных веществ (ацетилхолина, адреналина, се- ротонина, малых пептидов — гастрина и др.) в различные нервные структуры с помощью хемотродов, в сочетании с выработкой услов ных рефлексов.
-
Электрофизиологические методы, осно ванные на регистрации ЭЭГ, вызванных по тенциалов, нейронной активности различных отделов коры большого мозга, подкорковых структур.
-
Методы молекулярной биологии, на правленные на изучение роли молекул ДНК, РНК, других биологически активных веществ в образовании условных рефлексов.
-
Методы электронной микроскопии.
19.2.3. КЛАССИФИКАЦИЯ УСЛОВНЫХ РЕФЛЕКСОВ
В настоящее время не существует общепринятой научно обоснованной классификации условных рефлексов. Предлагается один из вариантов.
А. Все условные рефлексы подразделяют на те же группы, что и безусловные, на базе которых они были выработаны.
-
По биологическому значению различают рефлексы пищевые, оборонительные и др.
-
По виду рецепторов, на которые воздей ствует условный сигнал, различают экстеро- цептивные, проприоцептивные, интероцеп- тивньге условные рефлексы. Исследованиями лабораторий К.М.Быкова, В.Н.Черниговско го была показана связь коры большого мозга со всеми внутренними органами. Интероцеп- тивные условные рефлексы сопровождаются, как правило, расплывчатыми ощущениями, которые . И.М.Сеченов называл «темными чувствами», влияющими на настроение, ра ботоспособность. Проприоцептивные услов ные рефлексы лежат в основе научения чело века и животных двигательным навыкам (ходьба, производственные операции и др.). Экстероцептивные условные рефлексы фор мируют приспособительное поведение жи вотных по добыванию пищи, избеганию вредных сигналов, продолжению рода и т.д.
У человека ведущее значение имеют экстероцептивные словесные раздражители, формирующие сложные формы целенаправленного поведения.
3. По отделу нервной системы и характеру эфферентного ответа различают условные рефлексы соматические (двигательные) и вегетативные (сердечно-сосудистые, секреторные, выделительные и др.). Последнюю группу рефлексов подробно описал ученик И.П.Павлова К.М.Быков.
Желудок собаки наполняли водой через фистулу Басова так, чтобы животное не видело процедуры подготовки к вливанию. Эта процедура побуждает почки к усилению деятельности. Если вливанию воды в желудок предшествовал индифферентный сигнал, то через несколько подобных сочетаний вырабатывался условный рефлекс на деятельность почек: условный сигнал без вливания воды вызывал усиление ее работы в виде повышения мочеотделения (диурез). Данный условный рефлекс запускается при возбуждении действующих на экстероцепторы света или звука, но реализуется с помощью внутренних органов — почек.
Примером условнорефлекторного изменения деятельности внутренних органов могут служить условные рефлексы на фармакологические (лекарственные) средства. Введение последних в организм сопровождается определенными изменениями внутренних органов, что отражается на состоянии коры большого мозга, и это может служить подкреплением для действия индифферентной* сигнала. Например, если неоднократно сочетать звук метронома с введением апоморфина, вызывающего рвоту, то через некоторое время метроном сам по себе будет вызывать у человека рвоту без введения апоморфина. Это используется в качестве одного из методов лечения алкоголизма у человека. В этих случаях вводят апоморфин (о действии которого больной не догадывается). а затем через некоторое время, к моменту, когда должно возникать рвотное действие апоморфина, больному дают понюхать водку. У пациента формируется связь вида бутылки, запаха водки с пЪ-следующей рвотной реакцией. В клинических условиях манипуляции приготовления к введению или даче больному лекарства (вид человека, выполняющего лечебное воздействие, вид шприца) могут стать условными раздражителями. В клинической деятельности следует учитывать возможность образования условно рефлекторных связей на манипуляции врача, на лекарственные средства.
Существует ряд классификаций, применяемых только к условным рефлексам.
Б. По отношению сигнального раздражителя к безусловному (подкрепляющему) раздражителю все условные рефлексы делят на натуральные и искусственные (лабораторные).
1. Натуральные условные рефлексы формируются на сигналы, являющиеся естественными признаками подкрепляющего раздражителя. Например, запах, цвет мяса могут быть условными сигналами подкрепления мясом. Легко возникают условные рефлексы без специальной выработки на время. Например, прием пищи в одно и то же время ведет к выделению пищеварительных соков и других реакций организма (например, лейкоцитоз) к моменту приема пищи. Поскольку натуральные условные сигналы трудно измерить количественно (запах, цвет и др.), то в дальнейшем И.П.Павлов перешел к изучению искусственных рефлексов. Натуральные условные рефлексы отличаются большой прочностью и очень легко вырабатываются. "^'Искусственными (лабораторными) называют условные рефлексы на такие сигнальные раздражители, которые в природе не имеют отношения к безусловному (подкрепляющему) раздражителю. Основными из этих условных рефлексов являются следующие.
-
По сложности различают: а) простые ус ловные рефлексы, вырабатываемые на одиночные раздражители (классические условные рефлексы И.П.Павлова); б) ком плексные условные рефлексы — на не сколько сигналов, действующих одновре менно или последовательно; примером могут служить условные рефлексы пере ключения, заключающиеся в том, что один и тот же раздражитель может стать условным сигналом различных эффектор- ных реакций; в) цепные рефлексы — на цепь раздражителей, каждый из которых вызывает свой условный рефлекс; г) ин струментальные условные рефлексы.
-
По соотношению времени действия условно го и безусловного раздражителей различают рефлексы наличные и следовые. Для вы работки условных наличных рефлексов характерно совпадение действия условно го и безусловного раздражителей; послед ний включается лишь несколько позже. Следовые рефлексы вырабатывают в усло виях, когда безусловный раздражитель по дают через некоторое время после выклю чения условного, т.е. условный рефлекс вырабатывается на след от сигнального стимула.
-
По выработке условного рефлекса на базе другого условного рефлекса различают ус ловные рефлексы второго, третьего и дру гих порядков. Рефлексы первого поряд ка — это условные рефлексы, выработан ные на базе безусловных рефлексов (клас-
сические условные рефлексы). Условные рефлексы второго порядка вырабатываются на базе условных рефлексов первого порядка, при которых безусловный стимул отсутствует. Условный рефлекс третьего порядка образуется на базе условного рефлекса второго порядка. Чем выше порядок условных рефлексов, тем труднее идет их выработка. У собак удается образовать условные рефлексы только до третьего порядка.
• Условные рефлексы на время могут быть не только натуральными, но и искусственными. При неоднократной подаче безусловного раздражителя с неизменным интервалом между его подачами образуется рефлекс на время, т.е. за некоторое время до подачи подкрепления возникает условная эффекторная реакция.
В. В зависимости от сигнальной системы
различают условные рефлексы на сигналы первой и второй сигнальных систем. Последние вырабатываются только у человека: например, после образования условного зрачкового рефлекса на свет (сужение зрачка) произнесение слова «свет» также вызывает сужение зрачка у испытуемого.
Биологическое значение условных рефлексов состоит в их предупредительной, сигнальной роли: они имеют для организма приспособительное значение, готовя организм к будущей полезной поведенческой деятельности и помогая ему избежать вредных воздействий, тонко и эффективно адаптироваться к окружающей природной и социальной среде.
19.2.4. МЕХАНИЗМ ОБРАЗОВАНИЯ УСЛОВНЫХ РЕФЛЕКСОВ
Согласно И.П.Павлову (1903), временная связь образуется между корковым центром безусловного рефлекса и корковым центром условного раздражителя, т.е. замыкательная связь образуется в коре большого мозга (см. рис. 19.2). Импульсы, вызываемые индифферентным (условным) сигналом с любого участка кожи и других органов чувств (ухо, глаз), по нервам поступают в кору большого мозга и обеспечивают там образование очага возбуждения. Если после индифферентного сигнала подать пищевое подкрепление, то возникает более мощный второй очаг возбуждения в коре большого мозга, к которому направляется ранее возникшее и рассеявшееся по коре возбуждение (правило рассеива-
511
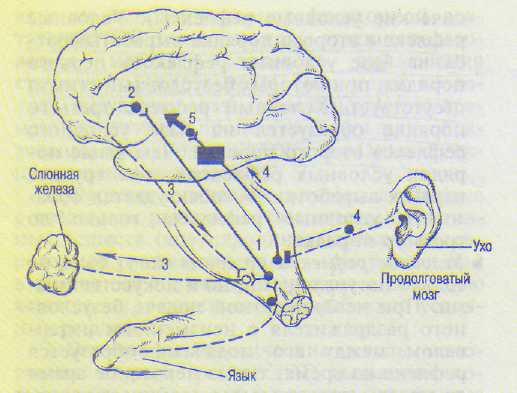
1 — афферентные волокна от вкусовых рецепторов; 2 — корковое представительство безусловного рефлекса; 3 — эфферентные волокна безусловного рефлекса; 4 — афферентные волокна от слуховых рецепторов; 5 — слуховая область коры. Жирная стрелка — установление временной связи.
ния возбуждения в мозговой коре). Это представление получило позже подтверждение в учении о доминанте (А.А.Ухтомский). Неоднократное сочетание в опытах индифферентного (условного) сигнала и безусловного раздражителя облегчает прохождение импульсов от коркового центра индифферентного сигнала к корковому представительству безусловного рефлекса (правило проторе: ния). И.П.Павлов назвал образование временной связи в коре большого мозга замыканием новой условнорефлекторной дуги: по-
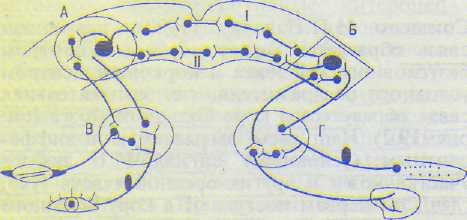
Рис. 19.3. Дуга условного рефлекса с двусторонней связью (по Э.А.Асратяну).
Л — корковый центр мигательного рефлекса, Б — корковый центр пищевого рефлекса; В и Г — подкорковые центры мигательного и пищевого рефлексов; I — прямая временная связь, II — обратная временная связь.
дача условного сигнала приводит к возбуждению коркового центра этого сигнала, оттуда импульсы направляются к корковому центру безусловного рефлекса и возбуждают его (рис. 19.2). В последние годы жизни И.П.Павлов допускал мысль о двустороннем проведении возбуждения в замыкательной связи. Впоследствии этот вопрос был специально изучен и подтвержден в лаборатории Э.А.Асратяна (1970). Им было показано, что образование.условных рефлексов с прямыми и обратными связями является общей закономерностью высшей нервной деятельности.
Э.А.Асратян полагал, что индифферентный сигнал не может быть вообще индифферентным, так как в начале своего применения вызывает собственную реакцию в виде ориентировочно-исследовательского рефлекса, который по мере повторения ослабляется. Допускают, что замыкание происходит между центрами двух безусловных рефлексов — сигнального и подкрепляющего. Это подтвердилось в опытах, где изучали два безусловных рефлекса — мигательный рефлекс, вызываемый струей воздуха в глаз, и безусловный пищевой (см. рис. 19.3). При их сочетании выработался условный рефлекс, причем если подавали воздушную струю, возникал пищевой рефлекс, а при даче пищевого раздражителя отмечалось мигание. Следовательно, временная связь имеет двусторонний характер.
Формирование условных рефлексов осуществляется с помощью механизмов памяти.
19.2.5. ПАМЯТЬ
Память — способность организма приобретать, сохранять и воспроизводить в сознании информацию и навыки.
Память обеспечивает человеку возможность активной, созидательной деятельности в обществе. Стирание следов памяти также важно, так как в противном случае могла бы возникнуть информационная перегрузка мозга.
Память различают по формам восприятия информации (логически-смысловая и чувственно-образная — зрительная, слуховая, моторная); по уровням усвоения (воспроизводящая и облегчающая); по происхождению в филогенезе (инстинкты) и в онтогенезе (условные рефлексы); по длительности хранения информации.
По длительности хранения информации выделяют несколько вариантов памяти, причем во всех из них первым этапом является
512
[сенсорная (иконическая) память — доли секунды (мгновенная память), далее: а) пер-Внная память — несколько секунд, вторичная — от нескольких минут до нескольких \т, третичная — на протяжении всей жизни; §5) кратковременная память — десятки секунд, долговременная —- более минуты и на протяжении всей жизни; в) кратковременная (первичная) — десятки секунд, кратковременная (вторичная) — от нескольких минут ^до нескольких лет, долговременная — на протяжении всей жизни. В рамках этих классификаций существуют противоречия. Например, термин «сенсорная память» отражает ее происхождение, а термины «кратковременная» и «долговременная» память свидетельствуют о длительности хранении информации, где также есть противоречия.
Во-первых, сенсорную память необоснованно называть особым видом памяти, так как при этом нет воспроизведения в сознании информации или навыка (воспоминания), а есть продолжение ощущения после выключения раздражителя. Это — результат инерционности анализаторов (последействие), например ощущение прикосновения, которого уже нет. Главный критерий в определении памяти — воспоминание. «Сенсорная память» этому критерию не соответствует.
Во-вторых, все противоречия о длительности хранения информации можно исключить, положив в основу классификации не длительность хранения информации, а механизм — электрохимические процессы и структурные изменения в синапсах и нейронах ЦНС. Идея длительности хранения информации сохраняется — кратковременная, промежуточная и долговременная память.
А. Кратковременная память. Наличие кратковременной памяти было доказано в опытах немецкого психолога Г.Эббингауса (1885), проводившего оценку эффективности воспроизведения человеком рядов случайных цифр, букв, символов. Эббингаус впервые установил, что случайные последовательности цифр, букв, символов после однократного прочтения, прослушивания можно воспроизвести без ошибок в строго ограниченном количестве единиц. Память, обеспечивающая удержание и воспроизведение оперативной информации, получила название кратковременной. Ее объем составляет примерно 7±2 единицы. Основным свойством данной разновидности памяти является непродолжительность. Хранение информации в кратковременной памяти длится секунды, минуты.
Механизмы кратковременной памяти. Единой теории до сих пор не существует. За-
поминание любой информации начинается с развития элементарных процессов в нейронных комплексах головного мозга — выделение различных медиаторов в синапсах, возникновение постсинаптических потенциалов действия. Большую роль для выяснения механизмов памяти сыграло применение в лечебных целях электрошока — сильного и кратковременного электрического воздействия на головной мозг. К_ак выяснилось, после электрошока, так же как и после механической травмы мозга в результате сотрясения, ушиба, наблюдается ретроградная амнезия, проявляющаяся в том, что человек полностью забывает о событиях, предшествовавших электрическому или механическому воздействию на мозг. По длительности ретроградной амнезии можно получить представление о времени консолидации памяти — времени перехода кратковременной памяти в долговременную. Консолидация памяти начинается обычно через несколько минут, иногда через несколько десятков минут (в опытах на крысах, у которых после выработки условного рефлекса вызывали электрошок, приобретенный навык сохранялся, если электрошок проводился через 10 мин и более после начала обучения, при более раннем воздействии на мозг навык исчезал). За 1 ч от начала обучения консолидация выражена достаточно хорошо. Информация запоминается лучше, если она привлекла внимание, — мозг сам ее повторяет и лучше усваивает.
Наблюдения за развитием состояния ретроградной амнезии у человека привели к формулировке одной из первых гипотез о механизме кратковременной памяти — гипотезе реверберации (циркуляции) возбуждения по замкнутым цепочкам нейронов. Считают, что в замкнутых нейрональных цепочках циркуляция длится в течение минут, сохраняя информацию в виде кода последовательности импульсов, передающихся от нейрона к нейрону. Допускают, что пока циркуляция продолжается, сохраняется нейрональный след воздействия того или иного раздражителя на организм в прошлом. С позиций гипотезы реверберации легко объясняется феномен ретроградной амнезии. Полагают, что развитие амнезии при электрошоке, травмах головного мозга возникает вследствие грубого нарушения ритмической активности в цепочках взаимосвязанных нейронов.
Б. Промежуточная память (по длительности хранения информации) — это процесс перевода кратковременной памяти в долговременную (консолидация памяти), который продолжается минуты — часы. Следы кратко-
513
временной памяти становятся устойчивыми через 3—4 ч, но еще могут быть стерты экстремальными воздействиями (наркоз и др.). На данном этапе электрофизиологические процессы запускают с помощью вторых посредников биохимические реакции. Поэтому данный период переработки поступившей информации и запоминания ее следует назвать нейрохимической памятью.
В настоящее время предпринимаются попытки объяснить природу кратковременной памяти, не прибегая к представлениям о реверберации возбуждения в ЦНС, хотя данных об иных механизмах кратковременной памяти все еще недостаточно. Существующие представления о механизмах кратковременной памяти позволяют назвать ее электрофизиологической памятью.
В. Долговременная память. Ее основой являются структурные изменения в нейронах (синтез белка), длительность — часы—дни-месяцы—годы. Объем долговременной памяти практически безграничен. Долговременная память устойчива к мозговым нарушениям. Уже за 1 ч наблюдается хорошая консолидация памяти, причем информация запоминается лучше, если она привлекла внимание. — мозг сам ее повторяет и лучше усваивает.
Долговременная память формируется с помощью механизмов кратковременной и промежуточной, поэтому четкой границы между ними нет (синтез белка при поступлении информации в виде нервных импульсов и выделении соответствующих медиаторов начинается уже через несколько минут, иногда через несколько десятков минут).
На первом этапе становления долговременной памяти важную роль играют синап-тические процессы. Так, один из типов ионо-тропных глутаматных синапсов после периода кратковременной активности, сопровождающийся входом кальция в нейроны, сохраняет повышенную вероятность генерировать потенциал действия в течение дней, недель и даже месяцев, что является клеточной формой памяти. Это явление, открытое в последние годы, дало надежду понять ключевые механизмы работы мозга. Дело в том, что долговременную потенциацию можно вызвать in vitro, т.е. в изолированном мозге, что открывает возможность исследовать этот феномен на клеточно-молекулярном уровне. Сотни лабораторий во всем мире интенсивно изучают долговременную потенциацию в гиппо-кампе — структуре мозга, который отвечает за пространственную память человека.
Показано также, что запоминание сигнальных свойств индифферентного возбуж-
Л
514
дения при выработке условных рефлексов может быть результатом синаптического облегчения реакций нейрона на индифферентный стимул в ходе его сочетаний с безусловным подкреплением. Соответствующий феномен получил название гетеросинаптичес-кого облегчения. В чем оно конкретно проявляется?
Будем проводить электрическую стимуляцию афферентных проводников, конвергирующих к одному и тому же нейрону. При слабом раздражении первого проводника («условный» стимул) нейрон отвечает небольшим возбуждающим пост-синаптическим потенциалом (ВПСП). При сильном раздражении второго проводника («безусловный» стимул, подкрепление) нейрон отвечает ВПСП большой амплитуды, на фоне которого генерируются потенциалы действия. Как выяснилось, сочетание первого и второго воздействий в течение нескольких минут постепенно приводит к усилению амплитуды ВПСП на «условный» стимул и возникновению потенциалов действия, аналогичных тем, которые возникали в ответ на «безусловный» стимул. Облегчение ответов на «условный» стимул в таких опытах сохранялось 10—40 мин после отмены «безусловного» стимула, т.е. характеризовалось следовым эффектом. Последний можно было трактовать как проявление запоминания и информационно значимого (условного) раздражителя.
Наиболее впечатляющие результаты при
проведении описанных выше экспериментов удалось зафиксировать на нейронах гиппо-кампа, где следовые изменения синаптичес-кой активности в процессе переработки входной импульсации сохранялись на протяжении часов и даже дней.
Каковы причины, лежащие в основе гете-росинаптического облегчения? Допускается следующий принципиальный механизм. Си-наптический контакт на некотором интегра-торном нейроне проводит возбуждение от условного раздражителя (рис. 19.4). Исходно нейрон на условный раздражитель не реагирует из-за низкой реактивности постсинап-тической мембраны синапса 1 к выделяемому медиатору 1. Однако под влиянием квантов медиатора здесь возникают кратковременные конформационные перестройки постсинап-тической мембраны. Синаптический контакт 2 обеспечивает передачу возбуждения на ин-теграторный нейрон от безусловного раздражителя. Выделяемый здесь медиатор 2 вызывает выраженную ответную реакцию. Предполагают, что возбуждения безусловного происхождения, помимо чисто электрических эффектов возбуждения нейрона-мишени, ускоряют протоплазматические биохими-
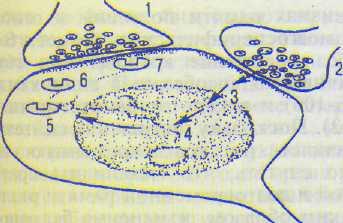
Рис. 19.4. Механизм повышения эффективности синаптической передачи в процессе образования временной связи между условным и безусловным возбуждением в ходе выработки условного рефлекса (по Х.Матиссу). 1—7 — описание в тексте.
ческие реакции (3), направленные на активацию генома (4) ядра нервной клетки. Результатами такой активации являются изменение синтеза РНК и синтез в рибосомах нейрона (5) нейроспецифического белка. В последующем вновь синтезированный белок (6) перемещается-в сторону постсинаптической мембраны того синапса, который подвергся под-пороговой активации в результате воздействия на него возбуждения условного происхождения. На заключительном этапе происходит встраивание белковой молекулы в постсинаптическую мембрану данного синапса, при этом функциональная активность синапса 1 изменяется: из неэффективного синапс превращается в эффективный.
Интересная гипотеза, объясняющая процесс трансформации функциональных свойств неэффективных синапсов в эффективные применительно к механизмам памяти, была предложена И.П.Ашмариным (1975). Допускается, что в результате непрерывного поступления к нейрону сигнальной информации в протоплазме нейрона усиливается синтез специфических белков-антигенов (глико-протеинов памяти), характерных для данного нейрона. Эти белки являются компонентами синаптических мембран, и, когда нейрон пребывает в состоянии относительного покоя, они синтезируются в количествах, достаточных только для их обновления. В период повторной импульсации, например, в ходе реверберации, при выработке ассоциативной связи происходит усиление синтеза белков-антигенов. В результате избыточного образования белки-антигены начинают перемещаться за пределы нейрона — в околонейронное (околосинаптическое) пространство. Воздействуя на расположенные вблизи нейрона клетки астроцитарной глии, белки-
антигены индуцируют процесс образования антител или антителоподобных субстанций. Антитела или антителоподобные субстанции вступают во взаимодействие с гликопротеи-нами в составе постсинаптических мембран тех же нейронов. Связываясь с ними, они видоизменяют функциональную активность синапса, при этом синапс из неэффективного переходит в разряд эффективных, начиная «узнавать» первичный раздражитель, обеспечивая его облегченное проведение через нейрон.
Важную роль в консолидации памяти играют нейропептиды. Показано, что пептиды могут находиться в пресинаптических терми-иалях в качестве сопутствующего медиатора. Например, вместе с норадреналином часто выделяются нейропептид У, опиоидные пептиды, соматостатин. Дофамин часто выделяется окончаниями аксонов вместе с холецис-токинином, энкефалином; ацетилхолин — с вазоактивным интестинальным пептидом, энкефалином, люлиберином; серотонин — с веществом П, тиролиберином, холецистоки-нином (И.П.Ашмарин). Доказано, что выделение пептидов в пресинаптических окончаниях зависит от частоты работы нейрона, при этом избыточное выделение пептида-спутника всегда наблюдается при усилении активности нейронов. Различные медиаторы могут оказывать разные эффекты в процессах усвоения и хранения информации. Серотонин, например, ускоряет обучение и удлиняет сохранение навыков при положительном эмоциональном подкреплении (например, пищевом). Норадреналин ускоряет обучение при отрицательном эмоциональном подкреплении (например, электрокожном). Как влияют пептиды на клетку-мишень? Пептид-спутник может значительно повысить сродство рецептора постсинаптической мембраны к основному медиатору. Например, вазоактивный интистинальный пептид (ВИП) усиливает сродство к ацетилхолину более чем в 10 000 раз. Пептиды выделяются нервными клетками не только в синаптическую щель, но и во внеклеточное пространство. Эффективный путь диффузии пептидов может составить 1 мм и более. Этого вполне достаточно, чтобы охватить своим влиянием (непосредственно или через систему вторых посредников — циклических нуклеотидов, ионов кальция) микроансамбль нейронов, изменяя его суммарную возбудимость, синхронизируя реактивность его нейронов, обеспечивая тем самым их включение в некоторый адаптивный поведенческий акт. Допускается, что мозаика образующихся функциональных
515
групп нейронов, подвергнутых нейрохимическому воздействию пептидов, может быть одним из механизмов оперативного функционального объединения нервных клеток в ходе обучения, в явлениях памяти.
Прямые доказательства участия пептидов в механизмах памяти можно получить, контролируя их выделение во внеклеточное пространство по показателям изменения химического состава цереброспинальной жидкости у животных при обучении. Несомненный интерес представляют опыты с АКТГ, мела-ноцитстимулирующим гормоном, вазопрес-сином. Как выяснилось, они обладают способностью стимулировать запоминание при введении извне (И.П.Ашмарин, Р.И.Крупников). Заметно улучшают обучение и память эндогенные опиатные пептиды — эндорфи-ны и энкефалиньг.
После открытия способа кодирования генетической информации в ДНК (генетической памяти) и успешного изучения иммунологической памяти были предприняты попытки обнаружить молекулярные основы нейронной памяти — возможного нервного субстрата энграммы. В частности, открытие структурных изменений нейронов в ходе формирования энграммы дало основание предполагать ключевую роль синтеза белка в консолидации памяти, т.е. в переводе информации из кратковременной в долговременную форму ее хранения.
Известна молекулярная гипотеза памяти П.К.Анохина (1968), согласно которой биохимические процессы, протекающие на уровне протоплазмы, вызывают динамические изменения генома нейрона, вызывая перестройку кода РНК, следствием чего является синтез адекватных для данной ситуации новых молекул белка, являющихся хранителем полученной информации. Допускают, что белковая молекула, в структуру которой вводится следовой отпечаток изменений молекул РНК, становится чувствительной к специфическому «узору» входного импульсного потока, что обеспечивает процесс его идентификации — узнавания (Х.Хиден).
Хотя механизмы долговременной памяти изучены недостаточно, гипотезы, акцент в которых делается на изменениях белкового обмена нейрона в качестве обязательного условия запоминания (консолидации памяти), в целом подтверждаются. Так, при угнетении механизмов, регулирующих синтез нейроспе-цифических белков, выработанные условные рефлексы при простых формах обучения сохраняются лишь на протяжении нескольких минут. Несомненные данные об участии в
механизмах памяти получены в отношении двух мозгоспецифических белков: S-100 и 14-3-2, содержание которых при обучении увеличивается, особенно в гиппокампе (белок S-100) и в коре головного мозга (белок 14-3-2). Поскольку подавление синтеза белка не влияет на кратковременную память, можно считать, что механизмы кратковременной и долговременной памяти различны.
Таким образом, изменение белкового метаболизма нейрона является решающим звеном сложнейшей цепи процессов формирования и закрепления следов памяти — энграммы. Следовательно, долговременную память можно называть структурной памятью.
Г. Роль отдельных структур головного мозга в формировании памяти. В ходе обучения запоминание осуществляется с помошью различных структур мозга, включающих два уровня: 1) неспецифический (общемозговой) — стволовая ретикулярная формация, гипоталамус, ассоциативный таламус, гиппокамп и лобная кора; 2) модально-специфический (региональный) — различные отделы новой коры большого мозга, за исключением лобной коры. Мозговая кора — основной субстрат модуляции памяти. Необходимо отметить, что разрушение корковых структур может вызвать расстройство памяти за счет нарушения разных процессов: либо запоминания, либо сохранения, либо воспроизведения. Височная кора участвует в запечатлении и хранении образной информации. При нарушении работы лобных долей затрудняется организация действий, отмечается повышенная отвлекаемость, наклонность к повторным стереотипным реакциям на предъявляемые раздражители.
Значительная роль в механизмах памяти отводится гиппокампу. Он является зоной конвергенции условных и безусловных стимулов, где мотивационное возбуждение гипоталамуса сопоставляется с информацией, поступающей из внешней среды через перегородку. Гиппокамп, с одной стороны, играет роль селективного входного фильтра, выделяя насущные стимулы, подлежащие хранению в долговременной памяти, устраняя реакции на посторонние для данного момента стимулы; с другой стороны, гиппокамп извлекает из памяти следы под влиянием моти-вационного возбуждения. Роль гиппокампа в обеспечении механизмов памяти иллюстрируется клиническим наблюдением эффектов, возникших после его двустороннего удаления по поводу эпилепсии. Описан случай, когда прооперированный больной потерял способность к запоминанию новой информации.
516
При этом не было обнаружено нарушений личности, каких-либо интеллектуальных расстройств, расстройств памяти на события минувших дней. Повторно больной был обследован спустя 28 лет (!) после операции. В отчете было отмечено: «Больной... не знает, где он живет, кто за ним ухаживает, что он ел в последний раз... В то же время у него сохраняются обрывки воспоминаний... к числу его любимых занятий относятся кроссворды и телевизор».
Ретикулярная формация оказывает активирующее влияние на структуры, участвующие в фиксации и воспроизведении следов памяти (энграмм). Она также включается в процессы формирования энграмм. Угнетение ее функций (например, нейрофармакологи-ческими средствами) сопровождается нарушением процесса консолидации памяти.
Таламокортикальная система способствует организации кратковременной памяти, при нарушении дорсомедиального и вентрояате-рального ядер таламической области затрудняются усвоение нового материала и сохранение ранее заученной информации. Влияние указанных ядер связывают с ослаблением их активирующего влияния на лобные отделы коры. В лобную кору основная информация поступает по двум путям: от сенсорных проекционных зон и через ассоциативные ядра таламуса. Оба потока импульсов в лобных долях обрабатываются и служат основой их специфической деятельности, связанной с формированием общих программ поведения и команд для ближайших подкорковых структур.
Хотя имеется специализация отдельных структур в запоминании, хранении информации, в выработке адаптивных навыков, мозг работает как единое целое. Об этом свидетельствуют, например, результаты опыта Г.Джаспера, в котором обезьяна обучалась передней конечностью выключать по условному сигналу электрический ток, оказывающий болевое воздействие на кожу. Если «обученную» конечность зафиксировать, процесс выключения тока по условному сигналу немедленно начинает обеспечиваться задней, т. е. «необученно й», конечностью.
Попытки локализовать энграмму в мозге млекопитающих дали два основных результата, которые лишь на первый взгляд кажутся противоречивыми. Во-первых, оказалось, что почти все корковые и подкорковые отделы головного мозга могут участвовать в запоминании, т.е. следы памяти не накапливаются в одной четко определенной его области. Наи-
более яркое свидетельство этого — сохранение у людей и животных даже с обширными повреждениями мозга удивительной способности к научению и запоминанию. Во-вторых, выявлено, что различные процессы памяти, вовлекая в этот процесс конкретную популяцию нейронов, не влияют на следы памяти, связанные с участием этой же популяции нейронов в формировании иной разновидности научения. Следовательно, механизмы памяти, реализуемые нейронными комплексами обширных областей мозга, в своем существенном проявлении являются механизмами исключительно высокоспецифичными.
19.2.6. ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
Изучение процессов торможения выявило две их формы — врожденные и приобретенные.
А. Врожденные формы торможения — внешнее и запредельное торможение.
1. Внешнее торможение — это торможение, которое проявляется в ослаблении или прекращении наличной условнорефлектор-ной реакции при действии какого-либо постороннего раздражителя. Например, если на фоне реализации условной реакции подать посторонний сигнал (звук, свет и др.) средней силы, то это вызывает появление ориентировочно-исследовательского рефлекса, который ослабляет или прекращает наличную условнорефлекторную деятельность. Этот рефлекс, возникающий на изменение внешней среды, рефлекс на новизну, И.П.Павлов называл рефлексом «что такое?». Он состоит в настораживании и подготовке организма на случай внезапно возникающей необходимости к действию. С повторением реакция на этот сигнал ослабевает и исчезает.
По характеру влияния посторонних раздражителей на условнорефлекторную деятельность они были разбиты на две группы — гаснущего тормоза и постоянного тормоза. Гаснущий тормоз — это посторонний сигнал, который с повторением его действия теряет свое тормозящее влияние, так как не имеет существенного значения для организма. В обыденной жизни на человека действует масса посторонних сигналов, на которые он сначала обращает внимание, а затем перестает их замечать. Постоянный тормоз — это такой посторонний раздражитель, который с повторением не теряет своего тормозящего действия. Сюда относят болевые раздражите-
517
ли, раздражения от переполненных внутренних органов (кишечника, мочевого пузыря и др.). Эти раздражители имеют существенное значение для человека и требуют от него принятия решительных мер к их устранению.
Механизм внешнего торможения. Согласно И.П.Павлову, посторонний сигнал сопровождается появлением в коре большого мозга нового очага возбуждения, вне дуги условного рефлекса (отсюда и название — внешнее торможение), который при средней силе раздражения оказывает угнетающее влияние на текущую условнореф-лекторную деятельность по механизму отрицательной индукции (доминанты). Внешне это похоже на реципрокное торможение. При этом имеет место срочное замещение одной целостной деятельности другой, более актуальной на данный момент.
2. Запредельное (охранительное) торможение возникает при действии на корковые клетки чрезвычайно сильных или длительных условных раздражителей. Между силой условного раздражителя и величиной ответной реакции имеется определенное соответствие («закон силы»): чем сильнее условный сигнал, тем сильнее условнорефлекторная реакция. Однако закон силы сохраняется до определенной величины, выше которой эффект начинает уменьшаться, несмотря на увеличение силы или частоты раздражения. При чрезмерной силе условного сигнала эффект его действия может полностью исчезнуть. Эти факты позволили И.П.Павлову утверждать, что корковые клетки имеют предел работоспособности. Поскольку появление указанного торможения не требует специальной выработки, оно по своей природе является безусловнорефлекторным.
Б. Приобретенное (внутреннее, или условное) торможение уловных рефлексов требует выработки. Отсюда название — условнореф-лекторное торможение. Согласно И.П.Павлову, оно локализовано в пределах («внутри») центральных структур условнорефлекторного акта. Внутренне заторможенные условные рефлексы называют также отрицательными условными рефлексами. Различают следующие виды условного торможения: угасатель-ное, дифференцировочное, условный тормоз и запаздывательное.
1. Угасательное торможение. В лаборатории И.П.Павлова было обнаружено, что повторное неподкрепление условных сигналов вызывает ослабление и устранение эффектов выработанного ранее положительного рефлекса. Через некоторое время он может восстановиться. Скорость угасания зависит от
интенсивности условного стимула и биологической значимости подкрепления: чем они значительнее, тем труднее совершается угасание положительного рефлекса. В обыденной жизни этот процесс связан с забыванием учебного материала, если он длительно не повторяется, устранением вредных привычек, навыков и др. Если во время действия условного угасательного рефлекса подействовать посторонним сигналом, возникает ориентировочно-исследовательский рефлекс, который ослабляет угасательное торможение и восстанавливает угасший ранее положительный рефлекс (явление растормажива-ния).
-
Дифференцировочное торможение раз вивается в случае неподкрепления раздражи телей, близких к подкрепляющему сигналу. Оно позволяет дифференцировать (разли чать) раздражители внешней среды. Напри мер, если у собаки тон 500 Гц подкреплять пищей, а тон 1000 Гц не подкреплять и их че редовать в течение одного опыта, то через не которое время животное начинает различать оба сигнала: на тон 500 Гц возникает поло жительная условная реакция в виде движе ния к кормушке, поедания корма, слюноот деления, а на тон 1000 Гц животное отвора чивается от кормушки с пишей, слюноотде ления не наблюдается, т.е. отмечается отри цательный дифференцировочный рефлекс. Таким способом можно добиться у животных выработки тонкого различения запахов, зву ков и др. Например, собаки-ищейки различа ют несколько тысяч запахов. У людей в осно ве различения окружающего мира, обучения, профессиональных навыков лежат процессы дифференцировочного торможения. Чем меньше различия между сигналами, тем труд нее идет выработка таких дифференцировоч- ных рефлексов. Удается выработать у живот ных различение частот метронома — 100 и 104 уд/мин, тонов 1000 и 995 Гц, распознава ние геометрических фигур, на раздражение различных участков кожи и др. Условное дифференцировочное торможение при дейст вии посторонних сигналов средней силы ослабевает и сопровождается явлением рас- тормаживания.
-
Запаздывательное торможение возни кает при отставлении подкрепления на 1 — 3 мин относительно начала действия услов ного сигнала. Постепенно появление поло жительной условной реакции сдвигается к моменту подкрепления. Более длительное от- ставление подкрепления у собак не удается. В выработанном условном запаздывательном рефлексе были выявлены две фазы — недея-
518
гельная и деятельная. Недеятельная фаза связана с образованием запаздывательного (внутреннего) торможения, так как этому торможению присуще явление растормажи-вания. Условный запаздывательный рефлекс необычен тем, что он одновременно является отрицательным (недеятельная фаза) и положительным (деятельная фаза) условным рефлексом. Выработка запаздывательного условного рефлекса наиболее трудна.
4. Условный тормоз возникает при добавлении к условному сигналу другого раздражителя и неподкреплении этой комбинации. Если условный сигнал подкреплять пищей, то вскоре вырабатывается положительный условный рефлекс. Например, условный сигнал «свет» подкрепляют, а комбинацию «свет и тон» не подкрепляют, превращая ее тем самым в условный дифференцировочный рефлекс. Теперь присоединение сигнала «свет» к любому положительному условному рефлексу усиливало последний, а присоединение «тона» — ослабляло. «Тон» стал уловным тормозом для любого условного рефлекса.
Посторонний раздражитель при действии условного тормоза вызывает растормажива-ние, что указывает на его отношение к внутреннему торможению.
Функциональные изменения при выработке положительных и отрицательных условных рефлексов (изменения возбудимости, ЭЭГ и др.) имеют общие черты, так же как и стадии формирования отрицательных рефлексов в общих чертах сходны с таковыми положительных рефлексов. Было показано, что образование отрицательных рефлексов не только устраняло замыкательную связь, ставшую неадекватной, но одновременно приводило к образованию другой замыкательной связи, противоположной по биологическому значению.
Значение всех видов внутреннего торможения условных рефлексов заключается в устранении ненужной в данное время деятельности — тонком приспособлении организма к окружающей среде.
Механизм внутреннего торможения условных рефлексов изучен недостаточно.