

1.2. Механизмы действия гормонов на клетки - мишени
Конечными эффектами действия гормонов на клеточном уровне могут быть изменения метаболизма, проницаемости мембраны для различных ве ществ (ионов, глюкозы и др.), процессов роста, дифференцировки и деления клеток, сократительной или секреторной активности и др. Реализация этих эффектов начинается со связывания гормона со специфическими клеточны ми белками-рецепт орами : мембранными или внутриклеточными (цито плазматическими и ядерными). Эффект действия гормонов через мембранные рецепторы проявляется относительно быстро (в течение нескольких минут), а через внутриклеточные рецепторы - медленно (от получаса и более).
Действие через м ем бранны е рецепторы типично для белково-пептид ных гормонов и производных аминокислот. Эти гормоны (за исключением тироидных), являются гидрофильными и не могут проникать сквозь били пидный слой плазмалеммы. Поэтому, гормональный сигнал передается в клетку по относительно длинной цепи, которая в общем случае выглядит так: гормон —> мембранный рецептор —> мембранный фермент —> вторичный посредник —> протеинкиназа —> внутриклеточные функциональные белки —> физиологический эффект.
Соответственно действие гормона через мембранные рецепторы реа лизуется в несколько этапов:
1 ) взаимодействие гормона с мембранным рецептором приводит к изменению конформации рецептора и его активированию;
2) рецептор активирует (реже - ингибирует) связанный с ним мем бранный фермент ;
3) фермент изменяет концентрацию в цитоплазме того или иного низкомолекулярного вещества - вторичного посредника',
4) вторичный посредник активирует определенную цитоплазматичес кую протеинкиназу - фермент, катализирующий фосфорилирование и из менение функциональных свойств белков;
5) протеинкиназа изменяет активность внутриклеточных функцио нальных белков, регулирующих внутриклеточные процессы (ферментов, ионных каналов, сократительных белков и т.п.), в результате чего реализу ется тот или иной конечный эффект действия гормона, например, ускоре ние синтеза или распада гликогена, запуск мышечного сокращения и др.
В настоящее время известны четыре типа ферментов, связанных с мембранными рецепторами гормонов, и пять основных вторичных посредников (рис. 1, табл. 1).
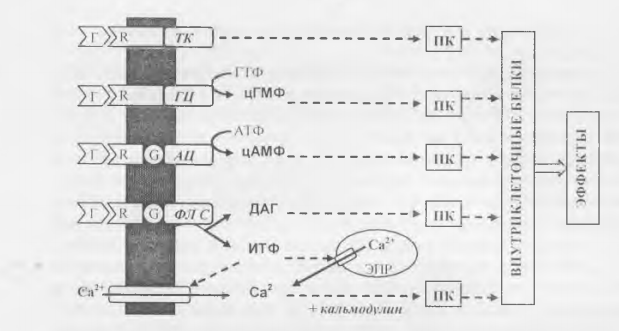
Рис. 1. Основные системы трансмембранной передачи гормонального сигнала.
Обозначения: Г - гормоны; R — мембранные рецепторы; G - G-белки; Ж - тирозин-
киназа; Г Ц - гуанилатциклаза; А Ц ~ аденилатциклаза; Ф.П С - фосфолипаза С; фл - мембранные фосфолипиды; ИТФ - инозитолтрифосфат, Д АТ - диацилглицерол; ЭПР - эндоплазматический ретикулум; ПК - различные протеинкиназы.
Таблица 1
Мембранные ферменты и вторичные посредники, опосредующие действие гормонов через мембранные рецепторы
|
Мембранный фермент |
G-белки |
Вторичные посредники |
Основные активирующие гормоны |
|
Тирозинкиназа |
Нет |
Нет |
инсулин, гормон роста, пролактин |
|
Гуанилатциклаза |
Нет |
цГМФ |
предсердный натрийуретический гормон |
|
Аденилатциклаза |
Есть |
цАМФ |
многие гормоны, например, адреналин через 3-адренорецепторы |
|
Фосфорилаза С |
есть |
ИТФ ДАТ Са2+ |
многие гормоны, например, адреналин через а г адренорецепторы |
В зависимости от того, как осуществляется связь между рецептороми мембранным ферментом различают два типа рецепторов: 1) каталитические рецепторы; 2)рецепторы, сопряженные с G-белками.
Каталитические рецепторы: рецептор и фермент связаны непосредственно (могут представлять собой одну молекулу с двумя функциональными участками). Мембранными ферментами у этих рецепторов могут быть:
• тирозинкиназа (разновидность протеинкиназ); действие гормонов через тирозинкиназные рецепторы не требует обязательного наличия вторичных посредников;
• гуанилатциклаза - катализирует образование вторичного посредника циклической ГМФ (цГМФ) из ГТФ.
Рецепторы, сопряженные с G-белками: сигнал от молекулы рецептора передается сначала особому мембранному G-белку1, который далее активирует или ингибирует определенный мембранный фермент, которым может быть:
• аденилатциклаза - катализирует образование вторичного посредника циклической АМФ (цАМФ) из АТФ;
• фосфолипаза С - катализирует образование из мембранных фосфолипидов двух вторичных посредников: инозитолтрифосфата (ИТФ) и диацилглицерола (ДАГ). ДАГ стимулирует протеинкиназу, а также является предшественником простагландинов и подобных им биологически активных веществ. Основной эффект ИТФ заключается в повышении содержания в цитоплазме еще одного вторичного посредника - ионов Са2+, которые поступают в цитозоль через ионные каналы плазмалеммы (из внеклеточной среды) или внутриклеточных депо Са2+(эндоплазматического ретикулума и митохондрий). Свое физиологическое действие ионы Са2+ осуществляют, как правило, в соединении с белком кальмодулином.
Действие через внутриклеточные рецепторы типично для стероидных и тироидных гормонов, которые благодаря своей жирорастворимости способны проникать через клеточные мембраны внутрь клетки и ее ядра (рис. 2).
Взаимодействуя с ядерными рецепторами, указанные гормоны влияют на процессы клеточного деления и реализации генетической информации (экспрессии генов), в частности, регулируют скорость биосинтеза функциональных клеточных белков - ферментов, рецепторов, пептидных гормонов и др.
В результате действия гормонов на цитоплазматические рецепторы изменяется активность клеточных органелл, например, интенсивность биологического окисления в митохондриях или синтеза белка в рибосомах.
В комплексе с цитоплазматическими рецепторами гормоны могут прони кать в ядро, действуя так же, как через ядерные рецепторы.
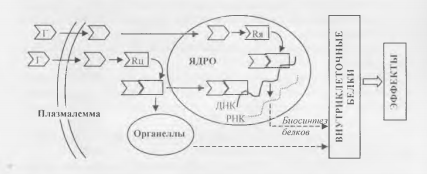
Рис.2. Механизмы внутриклеточного действия гормонов.
Обозначения: Г - гормоны; Rh - ядерные рецепторы; Rif - цитоплазматические рецепторы.